JBRA Assist. Reprod. 2019;23(4):352-360
ORIGINAL ARTICLE
doi: 10.5935/1518-0557.20190039
Expressed proteins and activated pathways in conditioned embryo culture media from IVF patients are diverse according to infertility factors
1Disciplina de Ginecologia Endocrinológica, Departamento de Ginecologia, Escola Paulista de Medicina da Universidade Federal de São Paulo (UNIFESP-EPM). Brasil
2Setor Integrado de Reprodução Humana, Departamento de Urologia, Escola Paulista de Medicina da Universidade Federal de São Paulo (UNIFESP-EPM). Brasil
3Huntington - Medicina Reprodutiva. Brasil
4Proteomics and Mass Spectrometry Facility, Center for Drug Discovery and Innovation (CDDI), University of South Florida (USF). USA
CONFLICT OF INTERESTS
The authors have no conflict of interest to declare.
ABSTRACT
Objective: Given that the embryo culture medium secretome reflects the embryo development, we hypothesize that protein profiles are affected according to infertility factors, which can be responsible for detrimental embryonic developmental competence. The aim of this study was to screen the protein profile of conditioned embryo culture media in patients presenting deep infiltrating endometriosis (ENDO) and polycystic ovarian syndrome (PCOS) undergoing IVF, by proteomics approaches. The control group was constituted by tubal factor patients.
Methods: Patients underwent in vitro fertilization (IVF) treatment as routine and oocytes were fertilized by ICSI. The embryos were group cultured until day 3 of development, and after transfer the culture media were collected. For the proteomics analysis, two pools of samples were prepared for groups CONTROL and PCOS, and 4 pools of samples for group DIE. Samples were prepared to deplete high abundant proteins and followed evaluated by high throughput proteomics approach.
Results: The embryonic organ and tissue development were physiological functions activated, based on proteins identified in the 3 study groups of samples. The samples coming from DIE patients presented a high calcium activity and on the other hand, embryos coming from PCOS patients showed a decreased calcium action. Other pathways as grow factors through the EGF signaling pathway overexpressed in ENDO culture medium and protein kinase A in PCOS were also observed.
Conclusions: Proteomic embryonic secretome will advance our knowledge of early embryogenesis and additionally could lead to improved selection of embryos for transfer warrants further investigation.
Keywords: embryo, conditioned culture media, secretome, IVF, endometriosis, ovarian polycystic ovarian syndrome
INTRODUCTION
In vitro fertilization (IVF) has gained attention since its introduction in 1978 and, to date, more than 2 million babies have been born worldwide through assisted reproductive technologies (ART). Developments in technologies for IVF, embryo culture and therapies for ovarian stimulation make ART relatively successful. However, it has its limitations with >50% of IVF embryos failing to implant. Selecting viable embryos for transfer is a key factor for the success of IVF treatment; and requires accurate pre-transfer assessment of embryo viability and morphology. However, the available methods to identify high implantation potential embryos are still limited (Gardner et al., 2015).
The field of human ART would therefore benefit from more quantitative methods of determining embryo viability and implantation potential to further improve the pregnancy rates. It is known that soluble ligands and its receptors mediate human pre-implantation embryo development and implantation (Thouas et al., 2015). Non-invasive analyses of the embryonic secretome, including proteins secreted by the embryo into the surrounding media, provide an alternative method for assessing an embryo's viability. These results may promote an understanding of the biological mechanisms, and potentially allow for the success of IVF developmental biomarkers.
A study evaluating conditioned media from non-manipulated human embryos cultured in vitro demonstrated that it contains extracellular vesicles and bearing the traditional microvesicle and exosome marker proteins CD63, CD9 and ALIX, which may suggested a way of communication at the maternal-fetal interface (Giacomini et al., 2017). A number of researchers has used the proteomics approach related to human reproduction in several subareas as ectopic pregnancy (Gerton et al., 2004), miscarriage (Liu et al., 2006), follicular fluid (Lo Turco et al., 2010; Kim et al., 2006; Silberstein et al., 2009; Estes et al., 2009; Twigt et al., 2012; Liu et al., 2007), oocytes (Ferreira et al., 2010), endometrium (Matorras et al., 2018) and embryos (Ferreira et al., 2010; Katz-Jaffe & Gardner, 2007; Katz-Jaffe et al., 2009; Dominguez et al., 2009). Recently, various proteins have been investigated as an embryo biomarker in the spent culture media (Butler et al., 2013; Mains et al., 2011; Ziebe et al., 2013; Cortezzi et al., 2013; Dominguez et al., 2008) given that the embryo culture medium secretome reflects the embryo development. We hypothesize that protein profiles are affected according with infertility factors, which can be responsible for detrimental embryonic developmental competence, ensuing on lower IVF success rates. Hence, we studied samples from patients presenting deep infiltrating endometriosis (ENDO) and polycystic ovarian syndrome (PCOS), which are the most frequent female factors of infertility. Endometriosis is present in until 50% (Missmer & Cramer, 2003) of infertile women and PCOS is present in the main cause of anovulatory infertility (Thessaloniki ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group, 2008). It is clear that the ovarian microenviroment is affected by the presence of endometriosis (Regiani et al., 2015; Singh et al., 2013; Karita et al., 2011) and PCOS (Huang et al., 2013; Ambekar et al., 2015; Roth et al., 2014)
Based on the knowledge that the potential of embryo development is an extension of oocyte quality, and the last is directly influenced by the microenviroment of ovary, possibly being affected by the presence of PCOS or ENDO, we hypothesized the embryo secretoma during its pre-implantation development could also vary reflecting the infertility factor condition. The aim of this study was to screen the protein profile of conditioned embryo culture media in patients with PCOS and ENDO undergoing IVF by proteomics approaches.
MATERIAL AND METHODS
Sampling
It was included in the study conditioned embryo culture media samples obtained from patients submitted to IVF cycle at the Huntington - Reproductive Medicine, Sao Paulo - Brazil. Institutional Ethical approval secured for the use of samples for the purpose of this research and the samples were donated by consenting patients whose signed the Informed Consenting Form as established by ethics for assisted reproduction treatment and research (CFM, 2013). Clinical characteristics and outcomes were obtained from patients' charts. All women presented with the following inclusion criteria: infertile patients undergoing ICSI cycles with ejaculated sperm, presence of both ovaries, regular menstrual cycle, body mass index (BMI) lower than 35 Kg/m2, no ongoing infectious disease, no uterus pathology, basal follicular stimulating hormone (FSH) measurement <14IU/L, and basal estradiol measurement <70pg/mL. The exclusion criteria were presence of gynecological bleeding, hydrosalpinx, allergy to gonadotropins or other medications used in the treatment, abusive use of any other medications during treatment, and male partners presenting with severe oligozoospermia. Among patients included, three groups were analyzed according to infertility etiology: tubal factor patients who were considered the control group for this study (CONTROL; n=6), infertile patients presenting PCOS (PCOS; n=7) and infertile patients presenting endometriosis grades III and IV (ENDO; n=14). The diagnosis of infertility was carried out according to international patterns. Patients were submitted to pituitary blockage and controlled ovarian stimulation as routine. The pituitary blockage was obtained with a GnRH agonist (Lupron kitTM, Abbot SA Societé Française des Laboratories, France), and the ovarian stimulation was performed using recombinant FSH (rFSH, Gonal-F ® Serono, Switzerland). When at least two follicles reached a diameter of 16 mm, the final follicular maturation was triggered with 250µg of recombinant hCG (rhCG, Ovidrel®, Serono, Switzerland). Oocyte retrieval was performed after 35 to 36 hours by transvaginal ultrasound-guided aspiration, and the luteal phase was supported by 90mg of daily progesterone (Crinone®, Serono, Switzerland) via vaginal approach. After oocyte recovery and denudation, all of the mature oocytes were fertilised by ICSI (Palermo et al., 1992) as per the routine of the clinic. The normally fertilized oocytes were identified and cultured in groups until day 3 (D3) in 1 mL of cell culture medium (G-1 Plus, Vitrolife) under a layer of paraffin oil (OVOIL, Vitrolife), in incubators with 5% O2 and 5% CO2. From D3 until the blastocyst stage (D5), the embryos were cultured in 1 mL of medium containing 10% human albumin (CSCM, Irvine Scientific) under a layer of paraffin oil in triple gas incubators (90% N2, 5% O2 and 5% CO2). The blastocysts were morphologically classified and the highest grades ones were selected for transfer to the women's uterus using a catheter guided by ultrasound. The culture media was collected on day 3 of development and stored at -80ºC until use. For the proteomics analysis, two pools of samples were prepared for groups CONTROL and PCOS, and four pools for group ENDO.
MS analysis
All analyzes in this step of study were carried out at Proteomics and Mass Spectrometry Facility, Center for Drug Discovery and Innovation (CDDI), University of South Florida (USF), Tampa - FL, USA. Arbitrary samples were used for methods standardization, and clinical samples were pooled into 3 to 4 samples pools according to pre-established groups (Control, PCOS and ENDO). Samples were prepared by using two steps of purification in order to eliminate the excess of albumin. First, samples were submitted to microfiltration using a 30KDa pore filter (Amicon® Ultra-0.5, Centrifugal Filter Devices - 30 kDa, Millipore, USA) and the filtrated material was collected. The material retained in the filter (higher than 30 KDa) was albumin depleted using affinity chromatography columns (Albumin & IgG Depletion SpinTrap, GE Healthcare Life Sciences, USA) according to manufacturer's protocol. The two aliquots of each sample were joined and protein concentrations were measured by Bradford method (Pierce 660nm Protein Assay Reagent, Thermo Scientific), using pre-diluted albumin standard curve (Pre-Diluted Protein Assay Standards: Bovine Serum Albumin, Thermo Scientific) and the absorbance's were acquired at 660 nm. Two hundred and fifty micrograms (250µg) of protein for each sample were diluted in 8M urea buffer and digested using FASP kits (Expedeon, Inc, USA) as per vendor specifications. Tryptic peptides were acidified by adding trifluoroacetic acid (TFA 1%) to get the 0.1% of TFA concentration, and then desalted on solid phase columns (DSC-18, Solid Phase Extraction - SPE, Discovery® DSC-18 SPE Products, Sigma Aldrich). Samples were vacuum dried (Wiśniewski et al., 2009). Tryptic peptides resulting from the preparation were fractionated by liquid chromatography mass spectrometry (HPLC-MS/MS). Five microliters injection of each sample loaded with aqueous solvent (0.1% formic acid in water) were separated by nano-flow reversed phase HPLC using a Nano-LC Ultra 2D+ (Eksigent, Dublin, CA) equipped with a Proteopep 2 Integra Fit trapping column (100 µm i.d. x 2.5 cm; C18, 5 µm, 300Ǻ) and a Proteopep 2 Integra Fit analytical column (75 µm i.d. x 10 cm; C18, 5 µm, 300Ǻ, New Objective, Woburn, MA). Samples (0.5 to 3 µg in 5 µL) were loaded onto the trap column at 2 µL/min (Solvent A) for 12 minutes, after which a valve was switched to include the analytical column. Peptides were then eluted with a gradient (300 nL/min) of 2% B to 35% B over 240 minutes (Solvent A: 100% H2O, 0.1% formic acid, Solvent B: 98% acetonitrile, 0.5% formic acid). Eluates were delivered by electrospray ionization (ESI) at 2 kV and analyzed by data-dependent MS/MS on a LTQ XL (Thermo Scientific, San Jose, CA, USA) mass spectrometer equipped with XCalibur (version 2.0.7) data acquisition software. Full MS scans were set for centroid mode at normal resolution. MS/MS scans were performed on the top ten most intense ions from each full scan to acquire spectral data for peptide identification. Dynamic exclusion durations were set to 180s with one repeat and a list size of 500. Raw data acquired were searched against the European Bioinformatics Institute's (EBI) universal protein resource database (UniProt, November 2014) using Mascot (version 2.2). Peptide and protein validations were performed using the Scaffold plataform (version 3.00.08). Search parameters used were as follows: precursor mass error tolerance of 20 ppm; fragment mass error tolerance of 0.1 Da; trypsin as a protease with one missed cleavage allowed; and carbamidomethylation of cysteine as fixed modification and oxidation of methionine as a variable modification. Identified peptides and proteins were validated and visualized with Scaffold 3.6 (Proteome Software, Portland, OR). The proteins identified in the previous analysis were analysed using IngenuityTM Pathway Analysis software (IPATM, QIAGEN, Redwood, USA). We considered peptide threshold of 20% and the biological processes were staggered according to the IPATM Knowledge Base. The association between the identified proteins and canonical pathways of the database was also accessed with IPATM software using Fisher's exact test (significance of p<0.01). From 2880 proteins identified, decoy proteins that are considered false identification were excluded. Two sets of analysis were carried out with remaining proteins using the IPATM. First of all we identified proteins exclusive in each study group (PCOS or ENDO) compared with CONTROL. We considered exclusively expressed those protein expressed in at least two sample pools of each group and absent on the other comparison groups. Then, we evaluated proteins differentially expressed in the study groups (PCOS or ENDO) compared with CONTROL and considered fold change higher than two.
RESULTS
Table 1 describes the demographic data regards the patients included in this study. In the ENDO group, 17 proteins were exclusively expressed (present in at least two of four samples pools), and two were over expressed compared to CONTROL (Table 2). The canonical pathways identified which were related to proteins exclusively expressed in ENDO group were associated with calcium metabolism [ calcium signaling (p=0.0057) and transport (p=0.0058) and calcium induced T -lymphocyte apoptosis (p=0.041) ] and EGF signaling (p=0.035) (Figure 1).
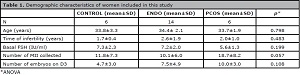
Table 1. Demographic characteristics of women included in this study

Table 2. Proteins exclusively expressed and over expressed in the ENDO compared to CONTROL group

Figure 1. Comparative analysis of canonical pathways based on proteins over-expressed in ENDO group compared to CONTROL.  CONTROL
CONTROL  ENDO.
ENDO.
On the other hand, in the PCOS group presented 284 proteins exclusively expressed and one overexpressed with fold change higher than 2 compared to CONTROL, which were associated with the following pathways: Protein Kinase A signaling and calcium signaling were downregulated, and GADD45 signaling, hydrocarbon receptor signaling and GDP-L fucose biosynthesis II were upregulated (Figure 2). The PCOS group had two pools of samples analysed and from 284 proteins exclusively expressed in the PCOS group, six were present in both pools of samples (Table 3).

Table 3. Proteins exclusively expressed and over expressed in the PCOS compared to CONTROL group
Based on proteins identified, the CONTROL group had the following cellular and molecular function highlighted: cellular development, cellular movement, amino acid metabolism, small molecule biochemistry, cellular assembly and organization, which were function associated to general cellular development. Also, the embryonic organ and tissue development were physiological functions activated based on proteins identified in the 3 study groups of samples.

Figure 2. Comparative analysis of canonical pathways based on proteins over-expressed in PCOS group compared to CONTROL. CONTROL PCOS.
DISCUSSION
In the last decade, the application of proteomics high throughput methodologies to human reproductive fluids and cells have delineated novel biochemical functional profiles and molecular processes that characterize and may affect folliculogenesis, oocyte maturation and quality, and as consequence the embryo development potential and clinical outcomes. Embryos are programmed to produce soluble ligands and receptors, which elicit changes in embryo developmental phenotype and also modulating local responses in the receptive endometrium determining an embryo maternal cross talk during peri-implantational period. These signaling pathways are highly complex and it understanding has been gradually improved (Thouas et al., 2015). The embryos from in vitro fertilization also produce soluble factors that are secreted in the spent culture media. Proteomics in spent culture media involved the measurement of amino acids (Brison et al., 2004; Sturmey et al., 2008), proteins (Katz-Jaffe et al., 2006a;b; 2009; Nyalwidhe et al., 2013) and metabolomics evaluates how the embryo alters its microenvironment (Scott et al., 2008; Leese et al., 2008). Authors have been trying to find if changes in the levels of some molecules are associated with implantation potential of embryos and clinical outcomes of IVF cycles.
Different approaches were necessary based on the nature of the specimens and the types of analyses being done. The standardization of proteomics approaches embryo culture medium was developed for this study, there we could obtain data available to a general protein profile in the samples. Depletion of abundant proteins combined with multidimensional protein fractionation was instrumental in allowing the study of middle- and lower-abundance proteins. Our study demonstrates that the technology can provide a consistent result given the restriction of starting material and time to analysis. These proteins with their regulatory pathways may play a vital role in reproductive process.
In embryos derived from ENDO group, we observed over expression of EGF signaling pathway compared to CONTROL. EGF is a growth factor family which has been already identified to be expressed by the human preimplantation embryo (Chia et al., 1995) and in vitro studies also showed the EGF treatment increases early human embryo development and blastocyst formation (Yu et al., 2012). Animal studies have shown that EGF enhanced developmental competence of cat embryos by stimulating cell proliferation and modulating the EGFR expression at various developmental stages (Thongkittidilok et al., 2015). On the other hand, an in vitro study cultured mice embryos with human tubal fluid from endometriosis patients and observed that the levels of embryonic EGF, IGF-I, and their receptors were increased, and it attenuated embryo development by impairing embryonic growth factor/receptor/signal transduction (Ding et al., 2010). Endometriosis has been associated with impaired IVF outcomes (Harb et al., 2013; Somigliana & Garcia-Velasco, 2015). Based on that, we can speculate the higher expression of EGF in endometriosis group may have an ovarian origin and it can be associated with the mechanism of embryo development of those patients.
Other pathways highlighted were associated with calcium signaling. Studies have shown that mitochondria has an enormous capacity to regulate Ca2+ (Giacomello et al., 2007). The embryo development is strongly correlated with the activity of mitochondria, and the abnormal distribution of mitochondria exert negative effects on the embryogenesis due to the abnormal ATP distribution (Nagai et al., 2006), because high energy supply around nucleus is very important during embryonic development (Wang et al., 2009), and may perform an important function in embryonic cell-cycle transition and embryonic axis establishment (Whitaker, 2008). A study as shown that conditioned medium from human embryos also trigger calcium oscillations in human endometrial epithelial cells, and importantly, the endometrial responses are affected by developmentally competency of embryos as low-quality human embryos trigger prolonged and disorganized calcium oscillations, leading to a uterine stress response (Brosens et al., 2014). In our study, while samples coming from endometriosis patients presented calcium-signaling upregulated, the PCOS samples showed it downregulated. In spite of most of publication find similar clinical outcomes after IVF in PCOS patients, it is related to alterations in oocyte quality and consequently in embryo quality, may be due to endocrine and intra-ovarian environment (Sermondade et al., 2013). The calcium metabolism downregulated in the PCOS embryos might reflect the poorer oocyte quality observed in those patients.
Protein kinase A signaling is also down regulated in PCOS samples. A study in non-human primate showed that mechanisms underlying adrenal hyperandrogenism seen in the human condition of PCOS are further considered in terms of the effects of altered relative expression of CYP17, HSD3B2 and CYB5, as well as the altered signaling responses of various kinases including protein kinase A (Abbott & Bird, 2009). GADD45, another protein over expressed in PCOS samples, is a protein that is often induced by DNA damage and other stress signals associated with growth arrest and apoptosis (Salvador et al., 2013).
The data on secretory activity can vary considerably suggest that embryo can intrinsically modulate the microenvironment and may reflect developmental plasticity rather than quality. In vitro studies provided evidences for a putative mechanism by which the decidualized stromal cells sense developmental embryos through the molecules secreted in the culture media (Brosens et al., 2014). Also, signals emanating from competent human embryos triggered a very specific transcriptional response in the mouse uterus, characterized by the induction of multiple metabolic genes (Teklenburg et al, 2010). Hence, focusing on exposures that infertility factors per se influence differential proteome profiles in the embryo culture media, can help understand its effects in the embryo developmental plasticity and competence, and consequently in IVF outcomes.
This study relies on the proteomic profile of conditioned embryo culture media in different infertility conditions. We did not correlated the proteins identified with the embryo characteristics as samples come from embryos cultured in groups and they were group cultured and we joined them into pools according to infertility factors to be analyzed. Also, the sample size is reduced and outcomes should be confirmed using immunodetection technique and in a higher number of samples. Due to high concentration of contaminants in the culture media, as albumin, samples were submitted to a number of process which might depleted other less abundant proteins. To date, the proteomic approach has proven to be a challenging task due to the complexity and diversity of the human embryo and heterogeneity across patients and within embryo cohorts.
In summary, the embryonic, organ and tissue development were physiological functions activated based on proteins identified in the three study groups of samples. The embryos coming from endometriosis patients present a high calcium activity and on the other hand, embryos coming from PCOS patients showed a decreased calcium action, which may be related to embryo developmental competence or plasticity. Other pathways as grow factors through the EGF signaling pathway overexpressed in endometriosis embryos and protein kinase A in PCOS were also observed. Characterizing the proteomic embryonic secretome will advance our knowledge of early embryogenesis and the embryo's role during the initial stages of implantation. Additionally, the activity in embryo culture medium could lead to improved selection of embryos for transfer warrants further investigation.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the assistance in sample collection of the IVF laboratory team in the Huntington Reproductive Medicine, São Paulo, Brazil, and the researchers at the Proteomics and Mass Spectrometry Facility, Center for Drug Discovery and Innovation (CDDI), University of South Florida (USF), Tampa - FL, USA, for their assistance with proteomics analysis.
They also thank Gabriela Venturini, PhD, University of Sao Paulo Brazil, for helping with bioinformatics analysis and Adriana Invitti, PhD, Federal University of São Paulo, for reading the manuscript and stimulating discussions.
Support
This study was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP; Grant numbers 2010/51877-1; 2010/51873-6) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPQ; Grant number 472930/2010-9).
ACKNOWLEDGMENTS
The authors gratefully acknowledge the assistance in sample collection of the IVF laboratory team in the Huntington Reproductive Medicine, São Paulo, Brazil, and the researchers at the Proteomics and Mass Spectrometry Facility, Center for Drug Discovery and Innovation (CDDI), University of South Florida (USF), Tampa – FL, USA, for their assistance with proteomics analysis. They also thank Gabriela Venturini, PhD, University of Sao Paulo Brazil, for helping with bioinformatics analysis and Adriana Invitti, PhD, Federal University of São Paulo, for reading the manuscript and stimulating discussions.
The abstract of this manuscript was presented as a poster at the 2018 European Society of Human Reproduction and Embryology (ESHRE) Annual Conference held in Barcelona during July 1-4, 2018.
REFERENCES
Abbott DH, Bird IM. Nonhuman primates as models for human adrenal androgen production: function and dysfunction. Rev Endocr Metab Disord. 2009;10:33-42.
Medline Crossref
Ambekar AS, Kelkar DS, Pinto SM, Sharma R, Hinduja I, Zaveri K, Pandey A, Prasad TS, Gowda H, Mukherjee S. Proteomics of follicular fluid from women with polycystic ovary syndrome suggests molecular defects in follicular development. J Clin Endocrinol Metab. 2015;100:744-53.
Medline Crossref
Brison DR, Houghton FD, Falconer D, Roberts SA, Hawkhead J, Humpherson PG, Lieberman BA, Leese HJ. Identification of viable embryos in IVF by non-invasive measurement of amino acid turnover. Hum Reprod. 2004;19:2319-24.
Medline Crossref
Brosens JJ, Salker MS, Teklenburg G, Nautiyal J, Salter S, Lucas ES, Steel JH, Christian M, Chan YW, Boomsma CM, Moore JD, Hartshorne GM, Sućurović S, Mulac-Jericevic B, Heijnen CJ, Quenby S, Koerkamp MJ, Holstege FC, Shmygol A, Macklon NS. Uterine selection of human embryos at implantation. Sci Rep. 2014;4:3894.
Medline Crossref
Butler SA, Luttoo J, Freire MO, Abban TK, Borrelli PT, Iles RK. Human chorionic gonadotropin (hCG) in the secretome of cultured embryos: hyperglycosylated hCG and hCG-free beta subunit are potential markers for infertility management and treatment. Reprod Sci. 2013;20:1038-45.
Medline Crossref
CFM-Conselho Federal de Medicina. Resolução CFM No 2.013/2013. Normas éticas para a utilização das técnicas de reprodução assistida. Brasília: CFM; 2013. Available at: http://www.portalmedico.org.br/resolucoes/cfm/2013/2013_2013.pdf
Chia CM, Winston RM, Handyside AH. EGF, TGF-alpha and EGFR expression in human preimplantation embryos. Development. 1995;121:299-307.
Medline
Cortezzi SS, Cabral EC, Trevisan MG, Ferreira CR, Setti AS, Braga DP, Figueira Rde C, Iaconelli A Jr, Eberlin MN, Borges E Jr. Prediction of embryo implantation potential by mass spectrometry fingerprinting of the culture medium. Reproduction. 2013;145:453-62.
Medline Crossref
Ding GL, Chen XJ, Luo Q, Dong MY, Wang N, Huang HF. Attenuated oocyte fertilization and embryo development associated with altered growth factor/signal transduction induced by endometriotic peritoneal fluid. Fertil Steril. 2010;93:2538-44.
Medline Crossref
Dominguez F, Gadea B, Esteban FJ, Horcajadas JA, Pellicer A, Simón C. Comparative protein-profile analysis of implanted versus non-implanted human blastocysts. Hum Reprod. 2008;23:1993-2000.
Medline Crossref
Dominguez F, Pellicer A, Simón C. The human embryo proteome. Reprod Sci. 2009;16:188-90.
Medline Crossref
Estes SJ, Ye B, Qiu W, Cramer D, Hornstein MD, Missmer SA. A proteomic analysis of IVF follicular fluid in women <or=32 years old. Fertil Steril. 2009;92:1569-78.
Medline Crossref
Ferreira CR, Saraiva SA, Catharino RR, Garcia JS, Gozzo FC, Sanvido GB, Santos LF, Lo Turco EG, Pontes JH, Basso AC, Bertolla RP, Sartori R, Guardieiro MM, Perecin F, Meirelles FV, Sangalli JR, Eberlin MN. Single embryo and oocyte lipid fingerprinting by mass spectrometry. J Lipid Res. 2010;51:1218-27.
Medline Crossref
Gardner DK, Meseguer M, Rubio C, Treff NR. Diagnosis of human preimplantation embryo viability. Hum Reprod Update. 2015;21:727-47.
Medline Crossref
Gerton GL, Fan XJ, Chittams J, Sammel M, Hummel A, Strauss JF, Barnhart K. A serum proteomics approach to the diagnosis of ectopic pregnancy. Ann N Y Acad Sci. 2004;1022:306-16.
Medline Crossref
Giacomello M, Drago I, Pizzo P, Pozzan T. Mitochondrial Ca2+ as a key regulator of cell life and death. Cell Death Differ. 2007;14:1267-74.
Medline Crossref
Giacomini E, Vago R, Sanchez AM, Podini P, Zarovni N, Murdica V, Rizzo R, Bortolotti D, Candiani M, Viganò P. Secretome of in vitro cultured human embryos contains extracellular vesicles that are uptaken by the maternal side. Sci Rep. 2017;7:5210. PMID: 28701751 DOI: 10.1038/s41598-017-05549-w
Medline Crossref
Harb HM, Gallos ID, Chu J, Harb M, Coomarasamy A. The effect of endometriosis on in vitro fertilisation outcome: a systematic review and meta-analysis. BJOG. 2013;120:1308-20.
Medline Crossref
Huang X, Hao C, Shen X, Liu X, Shan Y, Zhang Y, Chen L. Differences in the transcriptional profiles of human cumulus cells isolated from MI and MII oocytes of patients with polycystic ovary syndrome. Reproduction. 2013;145:597-608.
Medline Crossref
Karita M, Yamashita Y, Hayashi A, Yoshida Y, Hayashi M, Yamamoto H, Tanabe A, Terai Y, Ohmichi M. Does advanced-stage endometriosis affect the gene expression of estrogen and progesterone receptors in granulosa cells? Fertil Steril. 2011;95:889-94.
Medline Crossref
Katz-Jaffe MG, Gardner DK, Schoolcraft WB. Proteomic analysis of individual human embryos to identify novel biomarkers of development and viability. Fertil Steril. 2006a;85:101-7.
Medline Crossref
Katz-Jaffe MG, Schoolcraft WB, Gardner DK. Analysis of protein expression (secretome) by human and mouse preimplantation embryos. Fertil Steril. 2006b;86:678-85.
Medline Crossref
Katz-Jaffe MG, Gardner DK. Embryology in the era of proteomics. Theriogenology. 2007;68:S125-30.
Medline Crossref
Katz-Jaffe MG, McReynolds S, Gardner DK, Schoolcraft WB. The role of proteomics in defining the human embryonic secretome. Mol Hum Reprod. 2009;15:271-7.
Medline Crossref
Kim YS, Kim MS, Lee SH, Choi BC, Lim JM, Cha KY, Baek KH. Proteomic analysis of recurrent spontaneous abortion: Identification of an inadequately expressed set of proteins in human follicular fluid. Proteomics. 2006;6:3445-54.
Medline Crossref
Leese HJ, Baumann CG, Brison DR, McEvoy TG, Sturmey RG. Metabolism of the viable mammalian embryo: quietness revisited. Mol Hum Reprod. 2008;14:667-72.
Medline Crossref
Liu AX, Jin F, Zhang WW, Zhou TH, Zhou CY, Yao WM, Qian YL, Huang HF. Proteomic analysis on the alteration of protein expression in the placental villous tissue of early pregnancy loss. Biol Reprod. 2006;75:414-20.
Medline Crossref
Liu AX, Zhu YM, Luo Q, Wu YT, Gao HJ, Zhu XM, Xu CM, Huang HF. Specific peptide patterns of follicular fluids at different growth stages analyzed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Biochim Biophys Acta. 2007;1770:29-38. PMID: 16949757 DOI: 10.1016/j.bbagen.2006.06.017
Medline Crossref
Lo Turco EG, Souza GH, Garcia JS, Ferreira CR, Eberlin MN, Bertolla RP. Effect of endometriosis on the protein expression pattern of follicular fluid from patients submitted to controlled ovarian hyperstimulation for in vitro fertilization. Hum Reprod. 2010;25:1755-66.
Medline Crossref
Mains LM, Christenson L, Yang B, Sparks AE, Mathur S, Van Voorhis BJ. Identification of apolipoprotein A1 in the human embryonic secretome. Fertil Steril. 2011;96:422-7.e2.
Medline Crossref
Matorras R, Quevedo S, Corral B, Prieto B, Exposito A, Mendoza R, Rabanal A, Diaz-Nuñez M, Ferrando M, Elortza F, Ametzazurra A, Nagore D. Proteomic pattern of implantative human endometrial fluid in in vitro fertilization cycles. Arch Gynecol Obstet. 2018;297:1577-86.
Medline Crossref
Missmer SA, Cramer DW. The epidemiology of endometriosis. Obstet Gynecol Clin North Am. 2003;30:1-19, vii.
Medline Crossref
Nagai S, Mabuchi T, Hirata S, Shoda T, Kasai T, Yokota S, Shitara H, Yonekawa H, Hoshi K. Correlation of abnormal mitochondrial distribution in mouse oocytes with reduced developmental competence. Tohoku J Exp Med. 2006;210:137-44.
Medline Crossref
Nyalwidhe J, Burch T, Bocca S, Cazares L, Green-Mitchell S, Cooke M, Birdsall P, Basu G, Semmes OJ, Oehninger S. The search for biomarkers of human embryo developmental potential in IVF: a comprehensive proteomic approach. Mol Hum Reprod. 2013;19:250-63.
Medline Crossref
Palermo G, Joris H, Devroey P, Van Steirteghem AC. Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet. 1992;340:17-8.
Medline Crossref
Regiani T, Cordeiro FB, da Costa Ldo V, Salgueiro J, Cardozo K, Carvalho VM, Perkel KJ, Zylbersztejn DS, Cedenho AP, Lo Turco EG. Follicular fluid alterations in endometriosis: label-free proteomics by MS(E) as a functional tool for endometriosis. Syst Biol Reprod Med. 2015;61:263-76.
Medline Crossref
Roth LW, McCallie B, Alvero R, Schoolcraft WB, Minjarez D, Katz-Jaffe MG. Altered microRNA and gene expression in the follicular fluid of women with polycystic ovary syndrome. J Assist Reprod Genet. 2014;31:355-62.
Medline Crossref
Salvador JM, Brown-Clay JD, Fornace AJ Jr. Gadd45 in stress signaling, cell cycle control, and apoptosis. Adv Exp Med Biol. 2013;793:1-19.
Medline Crossref
Scott R, Seli E, Miller K, Sakkas D, Scott K, Burns DH. Noninvasive metabolomic profiling of human embryo culture media using Raman spectroscopy predicts embryonic reproductive potential: a prospective blinded pilot study. Fertil Steril. 2008;90:77-83.
Medline Crossref
Sermondade N, Dupont C, Massart P, Cédrin-Durnerin I, Lévy R, Sifer C. Impact of polycystic ovary syndrome on oocyte and embryo quality. Gynecol Obstet Fertil. 2013;41:27-30. Article in French.
Medline Crossref
Silberstein T, Saphier O, Paz-Tal O, Gonzalez L, Keefe DL, Trimarchi JR. Trace element concentrations in follicular fluid of small follicles differ from those in blood serum, and may represent long-term exposure. Fertil Steril. 2009;91:1771-4.
Medline Crossref
Teklenburg G, Salker M, Molokhia M, Lavery S, Trew G, Aojanepong T, Mardon HJ, Lokugamage AU, Rai R, Landles C, Roelen BA, Quenby S, Kuijk EW, Kavelaars A, Heijnen CJ, Regan L, Brosens JJ, Macklon NS. Natural selection of human embryos: decidualizing endometrial stromal cells serve as sensors of embryo quality upon implantation. PLoS One. 2010;5:e10258.
Medline Crossref
Thessaloniki ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Consensus on infertility treatment related to polycystic ovary syndrome. Hum Reprod. 2008;23:462-77.
Medline Crossref
Thongkittidilok C, Tharasanit T, Songsasen N, Sananmuang T, Buarpung S, Techakumphu M. Epidermal growth factor improves developmental competence and embryonic quality of singly cultured domestic cat embryos. J Reprod Dev. 2015;61:269-76.
Medline Crossref
Thouas GA, Dominguez F, Green MP, Vilella F, Simon C, Gardner DK. Soluble ligands and their receptors in human embryo development and implantation. Endocr Rev. 2015;36:92-130.
Medline Crossref
Twigt J, Steegers-Theunissen RP, Bezstarosti K, Demmers JA. Proteomic analysis of the microenvironment of developing oocytes. Proteomics. 2012;12:1463-71.
Medline Crossref
Wang LY, Wang DH, Zou XY, Xu CM. Mitochondrial functions on oocytes and preimplantation embryos. J Zhejiang Univ Sci B. 2009;10:483-92.
Medline Crossref
Whitaker M. Calcium signalling in early embryos. Philos Trans R Soc Lond B Biol Sci. 2008;363:1401-18.
Medline Crossref
Wiśniewski JR, Zougman A, Nagaraj N, Mann M. Universal sample preparation method for proteome analysis. Nat Methods. 2009;6:359-62.
Medline Crossref
Yu Y, Yan J, Li M, Yan L, Zhao Y, Lian Y, Li R, Liu P, Qiao J. Effects of combined epidermal growth factor, brain-derived neurotrophic factor and insulin-like growth factor-1 on human oocyte maturation and early fertilized and cloned embryo development. Hum Reprod. 2012;27:2146-59.
Medline Crossref
Ziebe S, Loft A, Povlsen BB, Erb K, Agerholm I, Aasted M, Gabrielsen A, Hnida C, Zobel DP, Munding B, Bendz SH, Robertson SA. A randomized clinical trial to evaluate the effect of granulocyte-macrophage colony-stimulating factor (GM-CSF) in embryo culture medium for in vitro fertilization. Fertil Steril. 2013;99:1600-9.
Medline Crossref