JBRA Assist. Reprod. 1997;01(03):117-125
ARTIGO DE ATUALIZAÇÃO
doi: 10.5935/1518-0557.1997.1.3.07
Desenvolvimento embrionário pré-implantação
Preimplantation embryo development
1Clínica ORIGEN: Centro de Tecnologia em Genética e Reprodução Humana
2Departamento de Cirurgia da Faculdade de Medicina da Universidade Federal de Minas Gerais
Resumo
Os autores descrevem detalhadamente os processos decorridos durante os primeiros dias de desenvolvimento embrionário, antes da implantação. Inicialmente, com a fertilização “in vivo” e “in vitro”, passando pelos estágios de clivagem, formação de mórula e blastocisto. Detalham também os processos de polarização dos blastõmeros, para melhor alocação celular intra-embrionária, compactação dos blastômeros, formação da blastocele e a primeira diferenciação celular, com formação de células de trofectoderma e embrioblasto, além da contagem do número de células, de ambos os grupos celulares.
Unitermos: embrião, pré-implantação, blastocisto, blastômero, FIV
Abstract
The authors describe in details the initial embryo development, over the first days before implantation. Initially, with "in vivo” and “in vitro” fertilization, through cleavage stages, mórula formation and blastocyst stage. Describe, in details, the polarization process reached by each blastomere within the embryo, compactation of the blastomeres and the first cell differentiation in trophectoderm cells and inner cell mass cells and also its cell number in both groups.
Keywords: embryo, preimplantation, blastocyst, blastomere, IVF
Introdução
Os gametas femininos ou oócitos encontram-se, em repouso, nos ovários, em estágio de prófase da primeira divisão meiótica. No período ovulatório, e sob ação do hormônio luteinizante, a meiose reinicia-se, originando o primeiro corpúsculo polar e, conseqüentemente, a liberação de metade do número de cromossomos. A seguir, a segunda divisão da meiose tem início, completando-se somente após a fertilização, com a expulsão do segundo corpúsculo polar. Os pronúcleos de ambos os gametas, contendo, cada um, metade do número de cromossomos, fundem-se em seguida, constituindo o zigoto, que, por sua vez, conterá o número completo de cromossomos (quarenta e seis). Este embrião original sofrerá várias divisões de clivagem, aumentando o número de células, que, paralelamente, terão seu tamanho reduzido.
Até recentemente, pouco se conhecia sobre o desenvolvimento embrionário humano, em seu período préimplantação, principalmente devido à inacessibilidade dos embriões humanos após a fertilização. Em estudo clássico de Hertig et al. (1954), foram realizadas descrições morfológicas a partir de limitado número de embriões, em estágio pré-implantação e após implantação inicial, obtidos de úteros removidos cirurgicamente. Com o desenvolvimento das técnicas de reprodução assistida para o tratamento de infertilidade, a fertilização de oócitos humanos “in vitro”, com posterior cultura de embriões até estágios anteriores à transferência ao útero materno (até 6 dias), tornou-se possível. A partir disto; embriões humanos em estágio de pré-implantação tornaram-se acessíveis para estudo.
O desenvolvimento de embriões de mamíferos, no período pré-implantação, tem sido classicamente dividido em três períodos. O primeiro período, denominado de zigoto, ou estágio de uma célula; estágio de clivagem, que dura até sua compactação, e o estágio de mórula, que finda com a formação do blastocisto. Este, por sua vez, é caracterizado pela primeira diferenciação celular, originando células do trofectoderma e do embrioblasto. O primeiro e fundamental requisito para estudarem-se os ciclos celulares e os tempos dos eventos, durante o desenvolvimento embrionário, é o estabelecimento de confiável ponto de referência. O momento da fertilização é, com freqüência, conveniente ponto a ser utilizado, principalmente quando se considerar a fertilização “in vitro” (Fig. I). Em seguida, pode-se identificar a presença de pronúcleos, aproximadamente 16 horas após a inseminação (dia 1). No dia 2, isto é, em torno de 48 horas após a inseminação, os embriões com desenvolvimento normal, possuem 2 a 4 células, e no dia 3, total de 6 a 10 células (Lewis & Wright, 1935). Até este estágio, os blastômeros têm aparência esférica, e com contornos visíveis. Entre os estágios de 16 e 32 células, os contornos celulares tornam-se menos distintos, devido à compactação, determinando a formação de mórula. No dia 5 após a inseminação, ocorre a formação dos blastocistos, tanto “in vitro” (Hardy et al., 1989), quanto “in vivo” (Buster et al., 1985). A implantação ocorre em torno de 6 dias após a fertilização, com as células do trofectoderma dando origem ao trofoblasto, enquanto as células do embrioblasto dão origem ao feto.
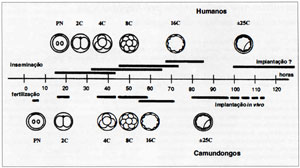
Figura I. Desenvolvimento embrionário no período pré-implantação.
Durante os primeiros 4 dias do desenvolvimento embrionário, o oócito recém-fertilizado sobrepõe várias divisões de clivagem, com partição da massa citoplasmática, até o estágio de 8 células, quando os blastômeros anteriormente esféricos, aparentemente maximizam seu contato uns com os outros de modo a formar agregado compacto, denominado mórula. De maneira diferente à ocorrida quando da divisão normal das demais células, não existe crescimento no que diz respeito ao aumento no volume citoplasmático. A divisão dos blastômeros filhos não é fenômeno sincrônico; entretanto, cada ciclo de divisões se faz completo antes que um novo se inicie, obedecendo os intervalos de 2, 4 e 8 células (Lewis & Wright, 1935; Colarco & Brown, 1969; Colarco & Epstein, 1973; Ducibella et al., 1977) (Fig. I).
O oócito fertilizado sofre 3 divisões de clivagem, determinando a formação de embrião de 8 células, o qual, pela primeira vez, apresenta células com elementos de formação epitelial, isto é, polarização e compactação (Ducibella & Anderson, 1975; Ducibella, 1977; Izquierdo et al., 1977). Após dois posteriores ciclos de divisão celular, o embrião atinge o estágio de blastocisto, constituído de população de células internas (embrioblasto) que originam o embrião, o feto e o neonato, e revestimento celular externo (trofectoderma), que tem por função transportar os metabólitos entre o embrioblasto e os tecidos maternos, iniciar a comunicação física e química com o útero materno e iniciar a formação placentária (Rossant, 1986; Pedersen, 1986). Além do advento da primeira diferenciação celular, uma barreira permeável é estabelecida, permitindo desenvolvimento subseqüente da cavidade blastocélica, por acúmulo de fluido interno. Inicialmente, pequenas cavidades repletas de fluido desenvolvem-se intercelularmente, entre os blastômeros (Benos & Balaban, 1980; Wiley, 1984). Estas pequenas cavidades fundem-se posteriormente, formando uma cavidade blastocélica única e maior. O revestimento epitelial e a cavidade intra-embriônica são as primeiras estruturas formadas durante a embriogênese nos mamíferos e sua subseqüente modificação estabelece a arquitetura tridimensional do embrião. A formação do embrioblasto e do trofectoderma no blastocisto, constitui a primeira e definitiva diferenciação celular, durante a embriogênese. Os dois tecidos demonstram diferenças em estrutura, função, bioquímica e potencial de desenvolvimento. A compreensão da origem destas sub-populações celulares e o mecanismo pelo qual elas são geradas durante o período de clivagem é ainda incerta. Algumas evidências supõem a hipótese de que o destino do desenvolvimento do blastômero em embrião de camundongo esteja relacionado à sua posição. Resultados de diversos experimentos indicam esta correlação na geração das células do embrioblasto e do trofectoderma, ainda em estágio de mórula (Mintz, 1964; Tarkowski & Wroblewska, 1967; Hillman et al., 1972; Handyside & Johnson, 1978). O mecanismo de compactação determina o surgimento de 3 principais características. A mais óbvia e evidente ao microscópio de dissecção é a mudança na forma individual dos blastômeros, passando de esférica e facilmente distinta, para a forma de cunha, achatando-se uns contra os demais, de modo a aumentar o contato celular. Este fenômeno é acompanhado pela formação de junções e desmossomos entre as células adjacentes (Ducibella et al., 1975; Ducibella & Anderson, 1979; Magnuson et al., 1978) além da reorganização que ocorre internamente em cada célula. O blastômero de embrião em estágio de 8 células, em fase inicial, apresenta-se esférico e sua superfície e organelas não aparentam nenhum sinal de orientação. O mesmo blastômero, entretanto, em fase tardia, apresenta sinais de polarização. As principais características do fenômeno da polarização são a reorganização da superfície celular, de modo que pequenos microvilos estejam localizados restritamente no pólo apical (Colarco & Epstein, 1973; Reeve & Ziomek, 1981) e a migração nuclear ocorra em direção à região basal do blastômero (Johnson & Ziomek, 1981). O citoplasma localizado entre o núcleo basal e o pólo apical, torna-se preenchido por diversas organelas (Kelly, 1981). Quando observados “in situ”, estes blastômeros têm seu eixo de polarização radiado a partir do centro do embrião (Fig II). Desta forma, os ápices celulares com os microvilos projetam-se externamente e o núcleo basal agrupa-se na região central (Reeve & Ziomek, 1981; Ducibella et al., 1977; Johnson & Ziomeck, 1981; Reeve, 1981) (Fig. II).
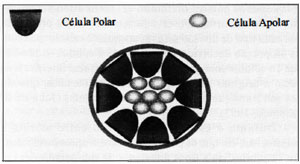
Figura II. Fenômeno de polarização celular.
Estudos de Ziomek & Johnson (1980) são bastante sugestivos no sentido de que o contato intercelular induz à polarização e que a assimetria deste contato determinaria o eixo da polaridade. Neste estudo, blastômeros isolados de embriões em estágio de 8 células tiveram baixa incidência de polarização; entretanto, quando colocados em contato com outro blastômero, ambos desenvolveram fenótipo polar. Mais ainda, o eixo de polaridade induzida em cada célula parecia ser determinado pelo ponto de contato intercelular.
A polaridade não é induzida meramente como conseqüência do acentuado grau de contato celular que ocorre durante a compressão das células no estágio de compactação (Pratt et al., 1982). De fato, a indução da polaridade precede a compressão celular (Ziomek & Johnson, 1980; Johnson & Ziomek, 1981; Reeve & Ziomek, 1981). Junções celulares especializadas são formadas inicialmente em estágio de 8 células. Estudos de Pratt et al. (1982) demonstraram evidências de que estas junções não teriam papel importante na formação ou transmissão de sinal para a polarização, ao impedirem o seu desenvolvimento, e confirmando que este impedimento não determinou a ação anti-polarizante. Na verdade, o exato mecanismo pelo qual a polaridade é induzida, permanece obscuro. O sinal indutor parece ser específico, regulado pelo desenvolvimento e não dependente de extensos contatos intercelulares ou junções especializadas.
Uma vez determinado o eixo de polaridade do blastômero, ele se torna estável. Estudos de Handyside (1980) e Ziomek & Johnson (1980) demonstraram que blastômeros, previamente induzidos à polarização “in situ”, mantiveram seu fenótipo polar mesmo após isolados. O eixo de polarização também é mantido estável durante a divisão celular. Segundo Johnson & Ziomek (1981) e Reeve (1981), blastômeros em estágio de 8 células, polarizados, identificados e analisados durante a divisão até 2 blastômeros, em estágio de 16 células, mantêm seus elementos de superfície e polaridade citoplasmática através do processo.
A diversidade celular surge, a partir das inferências de interações celulares, conseqüente polarização e da orientação dos planos de clivagem em blastômeros polarizados, que determinam se as divisões celulares devem obedecer a alguma orientação conservadora ou diferencial (Johnson et al. 1984). A divisão conservadora é caracterizada por partição do blastômero seguindo uma linha paralela ao seu eixo e determinando a formação de duas células polares com os pólos em contiguidade (Fig. III). Na divisão diferencial, a partição celularocorre segundo uma linha perpendicular ao eixo, sendo que uma das células incorpora a região apical, tornando-se, portanto, polar, enquanto a outra, derivada da região basal, é uma célula apolar (Fig. III). A freqüência em que as divisões conservadoras ou diferenciais ocorrem é variável e fator determinante da taxa de distribuição entre as células internas e externas do embrião de 16 células que, por sua vez, é responsável pela formação do embrioblasto e do trofectoderma (Johnson, 1989). Em estudos de Pedersen et al. (1986), Johnson (1986) e Fleming (1987), utilizando marcadores celulares, demonstraram que o número de células distribuídas para darem origem ao embrioblasto, pelo processo de divisão diferenciada, variou, entre embriões, de zero a 8, sendo que a maioria manteve a média de 5 células apolares em população de embriões de 16 células. Johnson & Ziomek (1981) e Handyside (1981) sugeriram que 7 em cada 8 divisões celulares em embriões de 8 células seria diferencial, sendo, a remanescente, conservadora. Isto determinaria a formação de um embrião de 16 células contendo 9 células polares e 7 células apolares. Segundo Johnson (1986), metade dos blastômeros do embrião de 8 células teria divisão conservadora e a outra metade, diferencial, originando um embrião de 16 células contendo 4 células apolares e 12 células polares.
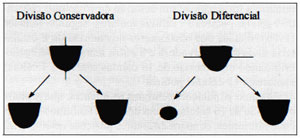
Figura III. Divisão celular em blastômeros polarizados.
As duas populações celulares resultantes, que formam o embrião de 16 células, além de terem fenótipos superficiais diferentes, ocupam diferentes posições internamente ao embrião e possuem diferentes propriedades. As células localizadas externamente possuem fenótipo polar, enquanto as células internas são apolares (Johnson & Ziomek, 1981; Handyside, 1981 e Ziomek & Johnson, 1981). Handyside (1981), utilizando marcadores celulares em embriões de 16 células intactos, corou as superfícies celulares expostas em vermelho. Após descompactação e desagregação embrionária em células únicas, as mesmas foram novamente coradas, utilizando-se novo marcador, que determinou predomínio da cor verde. A análise das células duplamente coradas demonstrou dois padrões diferentes. Algumas células não se coraram em vermelho, entretanto, apresentaram coloração uniforme em verde, indicando serem células internas e apolares do embrião original. As demais células apresentaram dupla coloração caracterizada por um polo em vermelho e o restante da superfície celular em verde. Estas células são, portanto, polares e possuem superfície exposta, constituindo as células externas do embrião intacto.
As células polares (externas) tendem a ser maiores que as células apolares (internas) e esta disparidade em tamanho pode ser decorrente da posição excêntrica do núcleo dos blastômeros polarizados, em estágio de 8 células. Além disso, as células polares possuem menor adesividade entre sí, do que as células apolares e menor grau de compressão. A diferença mais importante entre os dois tipos celulares é observada quando uma célula polar e outra apolar são agrupadas e colocadas em cultura. Segundo Ziomek & Johnson (1981), na maior parte dos casos, a célula polar emite um estreito processo citoplasmático que circunda a célula apolar, mantendo-a em sua recém formada cavidade, onde mais fluido pode ser acumulado subseqüentemente. Desta forma, as células polares mantêm sua posição externa ativamente e, mais ainda, confirmam a posição interna das células apolares.
As propriedades das células polares e apolares no embrião de 16 células (mórula), não apenas confirmam a estruturação de sua posição, como também representam características de diferenciação que antecipam as características do trofectoderma e embrioblasto, respectivamente. As células do trofectoderma são polares, envolvendo e transportando fluidos (Gardner & Johnson, 1972; Nadjica & Hillman, 1974; Burgoyne & Ducibella, 1977; Ducibella & Anderson, 1975), enquanto as células do embrioblasto são fortemente adesivas umas às outras e ocupam posição central (Gardner & Johnson, 1972; Rossant 1976; Johnson & Ziomek, 1982).
A segunda distribuição de células para a linhagem interna, ocorre na fase de 16 para 32 células, com os blastômeros polares dividindo-se, tanto diferencialmen-te, como de forma conservadora, sendo que a proporção de cada tipo de divisão será inversamente relacionada com a proporção decorrida no estágio de 8 células. Embriões de 16 células com grande número de células internas tendem a originar menor quantidade das mesmas, quando de sua transição para o estágio de 32 células (Johnson & Ziomek, 1983).
Durante o estágio de transição entre mórula e blastocisto, em que os embriões possuem aproximadamente 32 células, dois tipos diferentes de sub-populações celulares estão também presentes, localizados em diferentes posições (Nadjica & Hillman, 1974; Johnson & Ziomek, 1982). As células externas do trofectoderma são polares, transportadoras de fluidos e ligadas umas às outras por junções de alta complexidade, de modo semelhante às células externas dos embriões de 16 células.
O número médio de células externas em embriões, em estágio tardio de mórula, é de 20 e o de células internas gira em torno de 8. Nos recém-formados blastocistos, esta taxa estaria em torno de 18:14 (externas:internas) células (Handyside, 1978; Wiley & Eglitis, 1981; Johnson & Ziomek, 1982), o que está de acordo com o esperado para população de 9:7 (externas:internas) células em estágio de mórula.
Ziomek (1982) analisou as relações entre as linhagens celulares na mesma posição em estágio de 16 e 32 células, utilizando marcadores celulares individuais em estágio precoce e após seu desenvolvimento em blastocistos. Os resultados encontrados e também corroborados por outros autores (Randle, 1982; Geathurt et al., 1982), sugerem que, em embriões de 16 células, o destino das células apolares internas e das células polares externas são de fato o embrioblasto e o trofectoderma, respectivamente. Esta conclusão sustenta a teoria de que a polarização dos blastômeros de embriões de 8 células tem papel fundamental nas linhagens celulares do embrioblasto e trofectoderma.
Se considerarmos o destino dos blastômeros de embriões de 8 células como sendo fixo e determinado, a compreensão da teoria de totipotencialidade torna-se dificultada. Segundo estudo de Ziomek et al. (1982), dezesseis células polares, isoladas, de embriões de 16 células, e agregadas, em conjunto, formarão blastocistos contendo tanto células de embrioblasto, quanto do trofectoderma. O mesmo foi observado para células apolares. Estes resultados sugerem que a privação de qualquer célula de um embrião (seja ela polar ou apolar) determina desvio no curso de seu desenvolvimento natural, fazendo com que células polares originem células apolares para reposição e vice-versa.
Esta mudança de comportamento foi sugerida por Ziomek & Johnson (1981) e 1982), originada a partir da influência das interações celulares em seu plano de divisão. Células polares, comprimindo-se e circundando um núcleo de células apolares em embrião de 16 células, terão sua divisão realizada seguindo plano paralelo ou conservador, originando duas células polares. De modo semelhante, as células polares, completamente envoltas, darão origem a duas células apolares. Se as células polares forem privadas de células apolares, a compressão celular não será completa. Assim, as células polares com menor compressão, tenderão a dividir-se perpendicularmente, de modo diferencial, dando origem a uma célula polar e uma apolar. Este sistema teria função reguladora, isto é, quanto menor o número de células apolares, maior a possibilidade de se originarem mais destas células, de modo a manter relação embrioblasto: trofectoderma ajustada, para o caso de haver perdas ou morte celulares.
Com relação às células apolares, o contato assimétrico levaria à polarização, ou seja, ao colocarem-se células apolares por prolongado período na parte externa de um agregado celular, sua polarização seria induzida (Johnson & Ziomek, 1983). Novamente, a regulação interna da relação embrioblasto: trofectoderma é fornecida, isto é, a deficiência de células polares levaria a aumento de probabilidade de exposição de células apolares, determinando assim maior transformação destas em células polares.
A formação da blastocele, contendo fluidos, é característica marcante no desenvolvimento embrionário de mamíferos. O processo de formação de blastocistos envolve o rápido aumento no volume embrionário sem a necessidade do aumento de massa. Este aumento em volume parece facilitar o transporte do embrião através do trato genital, assim como a sua implantação. O processo de formação do blastocisto também converte uma estrutura tridimensional em um conjunto de camadas bidimensionais, nas quais ocorrerão as interações críticas que originarão a heterogenicidade celular (Barlow et al., 1972).
O primeiro epitélio diferenciado desenvolvido durante a embriogênense em mamíferos, é o trofectoderma: camada única de células polares de transporte, que envolve a cavidade blastocística. O trofectoderma desenvolve-se, concomitantemente, à produção de fluidos pela blastocele (cavitação), a partir do momento em que o embrião em estágio de mórula se desenvolve em blastocisto. É o único tecido capaz de efetivar a implantação e desenvolver-se em barreira imunológica, prevenindo a rejeição materna ao embrião.
O início da cavitação caracteriza-se por acúmulo de fluido entre os blastômeros quando o embrião encontra-se em estágio de 16 a 32 células. No ciclo de divisões posteriores, este fluido coalesce até formar uma única cavidade ou blastocele. O contínuo acúmulo de líquido determinará expansão no diâmetro do blastocisto até atingir o tamanho de blastocisto maduro. Neste ínterim, os blastômeros externos desenvolvem-se em epitélio escamoso (trofectoderma), que envolve a blastocele, enquanto os blastômeros internos formam o embrioblasto.
Com relação aos fatores determinantes do tempo de aparecimento da blastocele, não existe consenso. Estudos de Tarkowski & Wroblewska (1967) sobre a formação de blastocele em camundongos, após a remoção de blastômeros em estágio de 4 e 8 células, sugeriram que o número de células não é fundamental, pois após terem isolado um único blastômero, identificaram formação de blastocele em embrião contendo até 4 células. Estudo realizado por Snow (1973) demonstrou a formação de blastocele em embriões contendo apenas 2 células, após tratamento com citocalasina. Conclusões semelhantes foram descritas por Ducibella & Anderson (1975) e Granholm & Brenner (1976). Outras variáveis importantes seriam: o tempo decorrido desde a fertilização (idade cronológica do embrião); o número de divisões celulares; o número de replicações de DNA e cromossomos após a fertilização e a relação núcleo-citoplasmática. Em estudo de Smith & McLaren (1977), a análise da idade embrionária e o número de células na qual a cavidade blastocística iniciasua formação, indicam que o fator crítico pode ser determinado por alguma variável relacionada, mais ao número de células, do que à idade embrionária. Os experimentos em que se aumentava ou diminuía o número de blastômeros estabeleceram que a quantidade de células como fator isolado não estava envolvida no processo de formação do blastocisto, visto que os embriões que tinham seu número de células aumentado ou diminuído iniciaram sua formação de cavidade blastocélica, aproximadamente, no mesmo ponto de clivagem do grupo-controle, isto é, quando todas as células já tinham realizado uma replicação de DNA e, a maioria, se dividindo cinco vezes. O número de células, entretanto, era significantemente inferior ao esperado para o grupo-controle. Além disso, a supressão de uma divisão celular demonstrou que o número de células a partir da fertilização também não era fundamental. Os autores (Smith & McLaren, 1977) concluíram que o tempo de formação da blastocele seria determinado por algum fator, como o número de divisões nucleares ou replicação de DNA ou cromossomos, iniciados após a fertilização, ou mesmo a relação núcleo-citoplasmática.
Todos estes fatores são, entretanto, relativos às células individualmente, e não ao embrião como um todo. O que parece haver é alguma interligação direta entre eles, ou seja, a divisão nuclear coincide com a divisão celular, sendo que a replicação cromossômica ocorre um pouco mais precocemente e a síntese de DNA no meio do ciclo celular. A relação núcleo-citoplasmática tende a aumentar, uma vez que o conteúdo citoplasmático por genoma diplóide é dividido em dois a cada clivagem consecutiva. Este fato chama atenção para a hipótese de que o mecanismo do relógio biológico pode estar, de alguma forma, vinculado à relação núcleo-citoplasmática.
Alguns estudos mostraram evidências de que a relação núcleo-citoplasmática pode ser de importância para o desenvolvimento embrionário. Segundo Kaufman & Sachs (1976) e Modlinski (1980), a redução na relação núcleo-citoplasmática produzida por manipulação, é deletéria para o desenvolvimento precoce, inclusive pelo primeiro estágio de clivagem. Em estudo de McGrath & Solter (1983), a normalização, através de micromanipulação, da relação núcleo-citoplasmática permitiu, com sucesso, desenvolvimento em embriões de camundongos haplóides. Witkowska (1973) e Kaufman & Sachs (1976) observaram que, em embriões de camundongos ativados partogeneticamente (haplóides) e com relação núcleo-citoplasmática inferior ao normal, o aparecimento da blastocele foi retardado em até uma divisão de clivagem. Evsikov et al. (1990), utilizando embriões de camundongos, aumentaram e diminuíram a relação núcleo-citoplasmática através de micromanipulação e demonstraram que o fenômeno da cavitação não é dependente do número de divisões celulares, nem da relação núcleo-citoplasmática definitiva. Os autores sugerem que o início da cavitação seria dependente do tempo absoluto a partir do início do desenvolvimento.
Estudos relativos aos estágios tardios do desenvolvimento embrionário pré-implantação são bastante escassos, principalmente quando relacionados ao número de células do blastocisto e de sua primeira diferenciação celular, a qual determina a formação do embrioblasto e do trofectoderma. Em camundongos, a formação do blastocisto coincide com a quinta divisão de clivagem em estágio de 32 células, quando ocorre a diferenciação celular em trofectoderma e embrioblasto (Smith & McLaren, 1977; Handyside, 1978).
O número de células internas e externas, seja em estágio de mórula ou blastocisto, foi estudado através de reconstrução de embriões seccionados seriadamente (Barlow et al., 1972; Gardner & McLaren, 1974; Copp, 1978). Estes estudos, entretanto, são limitados devido ao pequeno número de embriões examinados, além da redução do número de células durante a fixação e da inabilidade em distinguir as células externas e internas, ao microscópio de dissecção (Handyside, 1981).
Utilizando-se técnicas de imunocirurgia, com a finalidade de lisar parcialmente as células do trofectoderma, é possível marcar diferencialmente os núcleos das células do trofectoderma e do embrioblasto, com uso de marcadores fluorocromos polinucleotídeos-específicos (Handyside & Hunter, 1984). Os núcleos do trofectoderma foram demarcados com um tipo de fluorocromo, durante a lise imunológica. Os embriões foram, então, fixados em etanol e os núcleos do embrioblasto corados com um segundo fluorocromo. Como o espectro de emissão de ambos os fluorocromos é diferente, os núcleos marcados podem ser distinguidos pela cor dos fluorocromos. Desta forma, não apenas se confirma a diferenciação celular em células do trofectoderma e do embrioblasto, como também pode-se contar especificamente o número de células de cada tipo presentes no blastocisto.
Handyside & Hunter (1986), utilizando as técnicas de marcação diferenciada das células do trofectoderma e embrioblasto de embriões de camundongos, observaram formação de blastocele coincidente com a quinta divisão de clivagem. Embriões estudados no dia 4, apresentaram 18,8 células do trofectoderma e 12,3 células do embrioblasto. No dia 5, o número total de células do blastocisto encontrado foi de 110 a 119, sendo 83 e 98 células do trofectoderma e 21 a 26, do embrioblasto. Este número de células é coincidente com o encontrado por Copp (1978), que estudou o desenvolvimento de embrião de camundongos “in vivo”, após secções seriadas.
Durante o período de 36 horas após a formação do blastocisto, o número de células do trofectoderma aumentou exponencialmente até atingir certo nível, o que coincide com as alterações celulares compatíveis com a implantação (Barlow et al., 1972). Em embriões humanos, Geber et al. (1992) observaram o fenômeno da divisão e diferenciação celular em células do trofectoderma “in vitro”, demonstrando aumento diário no número de células.
Estudos em embriões humanos, no período pré-implantação, são bastante raros. Após o desenvolvimento das técnicas de fertilização “in vitro” e cultura de embriões, antes de sua transferência para o útero materno, este período da vida embrionária foi mais facilmente compreendido. A compactação ocorre no dia 3 pós-fertilização “in vitro”, no estágio de 8 a 16 células, a blastulação, no dia 5 ou 6 e a implantação, “in vivo”, entre os dias 7 e 10. O tempo transcorrido para completar os três primeiros ciclos de clivagem é semelhante ao encontrado no camundongo, com média aproximada de 60 horas (Edwards et al., 1981; Sundstrom et al., 1981; Trounson et al., 1982). Para os estágios posteriores, existe pouca informação relativa ao número de células ou à sua diferenciação em trofectoderma ou embrioblasto no estágio de blastocisto. Hertig et al. (1954) recuperaram 2 blastocistos com idade aproximada de 5 dias pós-fertilização, em úteros removidos cirurgicamente. A identificação do número de células foi realizada através da contagem de núcleos em secções seriadas, sendo encontradas 58 e 107 células, sendo que 5 e 8 células, respectivamente, foram consideradas como relativas ao embrioblasto, baseando-se apenas em sua posição. Croxatto et al. (1972) obtiveram 1 blastocisto, a partir de lavagem uterina, que, após corte seriado, mostrou ter 185 células. Entretanto, este apresentava idade superior a 5 dias pós-fertilização.
Steptoe et al. (1971), contando os núcleos de 2 blastocistos desenvolvidos “in vitro” por período de 7 dias após a fertilização identificaram 110 e 112 núcleos.
Hardy et al. (1989) estudaram 149 embriões humanos normalmente fertilizados, que determinaram taxa de formação de blastocistos de 42% (63 embriões) no dia 5 ou 6 pós-fertilização. O número total de células, número de células do trofectoderma e do embrioblasto, foi analisado através de marcação diferenciada dos núcleos com fluorocromos polinucleotídeo-específicos. Os blastocistos récem-formados no dia 5 tinham o total de 58,3 células, aumentando para 84,4 e 125,5, nos dias 6 e 7, respectivamente. O número de células do trofectoderma foi similar nos dias 5 e 6 (38 e 40 respectivamente), dobrando no dia 7 (80,6). Em contraste, o número de células do embrioblasto, dobrou entre os dias 5 e 6 (20 e 42, respectivamente), mantendo-se virtualmente não modificado no dia 7 (46). O número mínimo de células observado em blastocistos morfologicamente normais, nos dias 5 e 6, foi de 24 e 27, respectivamente.
As taxas de formação de blastocisto de embriões humanos “in vitro” foram também pouco estudadas. No estudo supracitado de Hardy et al. (1989), a freqüência de formação de blastocisto encontrada foi de 42%. Hardy et al. (1989b) identificaram a formação de blastocistos em 58% de 43 embriões normalmente fertilizados (25 blastocistos) no dia 6, pós-fertilização “in vitro”. Lopata & Hay (1989), estudando embriões humanos em dois meios de cultura diferentes, identificaram formação de blastocistos de 26,5% (104/392) em meio mínimo essencial e 14,5% (34/234) em meio HTF (fluido tubário humano sintético). Bolton et al. (1989) demonstraram taxa de formação de blastocisto de 17% (55/317), pós-fertilização “in vitro”. Dokras et al. (1991), em estudo de embriões humanos pós fertilização “in vitro”, identificaram taxa de 14,5% (248/1708) de formação de blastocistos em embriões normalmente fertilizados. Geber et al. (1995) observaram taxa de formação de blastocisto de 53% (25/47), em embriões humanos biopsiados.
O fenômeno de extrusão do blastocisto da zona pelúcida (hatching) é essencial para a implantação do mesmo no útero materno. A zona pelúcida, camada glicoprotéica que reveste o oócito e embrião de mamíferos, é de importância funcional e estrutural durante os processos de fertilização e desenvolvimento pré-implantação. Durante a fertilização, em algumas espécies, a zona pelúcida é responsável pela indução da reação acrossômica (Bleil & Wassarman, 1980) e, prevenção e controle da polispermia (Yu & Wolf, 1981). A função da zona pelúcida, durante os estágios de clivagem embrionária, parece estar relacionada ao aspecto físico, isto é, prevenção da dispersão e perda dos blastômeros e impedimento de contato direto entre o embrião e células estranhas, como epitélio do trato reprodutor feminino, espermatozóides ou leucócitos (Bronson & McLaren, 1970; Modlinski, 1970). Além disso, a zona pelúcida facilita a passagem do embrião através das tubas uterinas. Estudos de Lindenberg et al. (1989) demonstraram que 25% dos embriões humanos em estágio de blastocistos, fertilizados “in vitro”, apresentavam extrusão da zona pelúcida. Hardy et al. (1989) identificaram taxa de extrusão de 10% (6/63) em blastocistos humanos. Lopata & Hay (1989) obtiveram 21,7% (30/138) de taxa de extrusão em blastocistos humanos cultivados em dois tipos distintos de meio de cultura. Dokras et al. (1991) observaram taxa de extrusão de 18,1% em blastocistos humanos fertilizados “in vitro”. Em embriões submetidos a micromanipulação, a taxa de extrusão apresenta-se aumentada por motivos óbvios, podendo atingir taxas de 48% dentre os blastocistos formados (Geber et al. 1995).