JBRA Assist. Reprod 2006;10(4):24-29
ARTIGO DE REVISÃO
doi: 10.5935/1518-0557.2006.10.4.05
Biologia Molecular na Espermatogênese
Molecular Biology in Spermatogenesis
1Instituto Valenciano de Infertilidad (IVI)
2FIVIER (Fundación IVI para el Estudio de la Reproducción)
ABSTRACT (PORTUGUÊS)
As técnicas de biologia molecular sofreram importantes avanços na ultima década e sua aplicação pratica é cada dia mais presente. Os estudos realizados envolvendo a gametogênese masculina mostram nítida correlação entre achados moleculares e quadros clínicos e fenotípicos. Alterações genéticas estão relacionadas com a infertilidade masculina como falha na espermatogênese, alterações de secreção de gonadotrofinas e ausência de ductos deferentes. O estudo molecular das células espermáticas mostra que alterações no empacotamento e na integridade do DNA podem levar a defeitos na formação do pro-núcleo masculino e, portanto, na performance da fertilização in vitro. Mostra também que existe aumento de anomalias no DNA e aumento de apoptose nos espermatozóides de amostras de sêmen congelado. A citometria de fluxo pode identificar populações de células espermáticas enriquecidas e permitir sua seleção antes de um tratamento de ICSI. Através destas técnicas se pôde desenvolver ou se desenvolverão os tratamentos paliativos ou de reprodução assistida necessários para que os casais nos quais os homens estejam afetados possam ter descendência sem correr riscos.
Palavra-chave: ESPERMATOGÊNESE, INFERTILIDADE MASCULINE, DNA, BIOLOGIA MOLECULAR
ABSTRACT
Molecular biology techniques have suffered important advances in the past decade and its practical applications are getting more present each day. The studies involving male gametogenesis show a subtle relationship between molecular findings and clinical features and fenotypes. Genetic alterations are related to male infertility like spermatogenesis failure, impaired gondotropin releasing and deferent duct agenesia. The molecular study of the spermatic cells shows that impairment of DNA packing and its integrity leads to defects in the pro-nucleus formation and, thus, interferes with in vitro fertilization outcome. These studies also show that there is an increase in DNA anomalies and increased apoptosis in spermatozoids from thawed semen. Flow cytometry can identify enriched cell populations and allow their selection before an ICSI treatment. Through these techniques, it will be possible to develop the palliative or assisted reproduction treatments needed for the couples where there is a male factor to achieve pregnancy without risks.
Key Words: Spermatogenesis, male infertility, DNA, Molecular Biology
INTRODUÇÃO
A espermatogênese é um processo biológico que pode resumir-se em três fases consecutivas: na primeira delas, ocorrem proliferação e renovação das espermatogônias, graças a mitoses sucessivas, que resultarão, por um lado em novas células madre e, por outro, em células que avançarão na espermatogênese. Estas darão lugar a espermatócitos, que entrarão em processo de diferenciação citológica que concluirão com a meiose, o que reduzirá sua carga genética dando espermátides haplóides. Finalmente, as espermátides entrarão no processo de espermiogênese, no qual é formado o acrossomo, a cauda e ocorre empacotamento do DNA, resultando finalmente no espermatozóide haplóide maduro.
Durante a década passada, as ferramentas da biologia molecular moderna proporcionaram descobertas únicas no conhecimento da espermatogênese.
A investigação citogenética em homens inférteis mostra que as anormalidades cromossômicas autossômicas e sexuais podem ser responsáveis por deficiências na espermatogênese. Junto com a infertilidade de origem cromossômica, as anomalias meióticas ou espermáticas sugerem a presença de mutações gênicas que interferem na evolução da espermatogênese e levam à formação de espermatozóides anormais. Através da Biologia Molecular se pode demonstrar a existência de essas mutações. Estas técnicas permitem que se conheça o defeito de células germinais portadoras de uma anomalia cromossômica particularmente quando esta é equilibrada e envolve autossomos, e não os cromossomos sexuais (Guichaoua & Luciani, 2003).
Existe necessidade de que ocorram mudanças na cromatina durante a espermatogênese, formando espermatozóides nuclearmente maduros que sejam capazes de fecundar um óvulo e dar lugar a um pronúcleo masculino. Essas mudanças podem ser analisadas pelo estudo do DNA.
A cromatina das células somáticas geralmente difere da cromatina dos espermatozóides, e esta diferença é baseada principalmente na ploidia, já que somente um jogo completo de cromossomos está incluído no núcleo de cada espermatozóide, enquanto existem duas cópias de cada cromossomo no núcleo das células somáticas. Além disso, outra diferença existente é o nível de empacotamento de cromatina dos espermatozóides, já que nestes ocorre uma mudança nas histonas de união do DNA por proteínas de transição que, por sua vez, serão substituídas por protaminas nas espermátides (Green et al., 1994).
Após a entrada de um espermatozóide em um ovócito, a nucleoplasmina presente no citoplasma do gameta feminino produzirá a separação das protaminas por meio da redução das pontes de dissulfeto estabelecidas com o ácido nucléico, fazendo com que ocorra a descondensação do núcleo e aumente seu tamanho, formando o pronúcleo masculino, que se combinará com o feminino no processo de fecundação (Philpott et al., 1991; Philpott et al., 1992).
Com essa informação podemos concluir que, em cada etapa da espermatogênese, existe uma situação concreta do DNA, extremamente controlada e que qualquer alteração poderia dificultar o processo reprodutivo.
CONTROLE GÊNICO DA ESPERMATOGÊNESE
Com o sequenciamento do genoma humano (International Human Genome Sequencing Consortium, 2001:( Venter et al., 2001)) e a implementação dos programas funcionais em grande escala, tem-se a oportunidade de identificar novos genes implicados na espermatogênese. Atualmente, somente as mutações pontuais no gene receptor de andrógenos (RA) representam um dos poucos defeitos gênicos realmente associados com uma falha espermatogênica. A realização de estudos em ratos knock out (onde se anula seletivamente a expressão de um determinado gene) mostra que muitos genes codificados tanto nos cromossomos sexuais como nos autossômicos estão implicados na diferenciação das células germinais, meiose ou nas etapas sucessivas da espermiogênese (Tabela I).
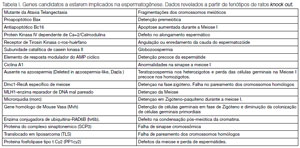
Tabela I. Genes candidatos a estarem implicados na espermatogênese. Dados revelados a partir de fenótipos de ratos knock out.
O estudo físico, genético e funcional do cromossomo Y permitiu especial atenção à região Yq11, onde parecem estar localizados alguns genes da espermatogênese (fatores de azoospermia “AZF”) (Maclahlan et al., 1998). Aproximadamente 15% dos homens em que se diagnostica uma oligozoospermia severa ou uma azoospermia são portadores de microdeleções nesta zona, por perda de segmentos específicos de DNA que levam à perda de genes vitais para a produção espermática (Chandley et al., 1998). Concretamente foram localizadas três regiões determinadas AZFa, AZFb e AZFc (Quershi et al., 1996; Vogt et al., 1996). Dos genes, o DAZ (ausente na azoospermia) e o RBM1 (domínio de união ao RNA) codificam proteínas de união ao RNA e se acredita que desempenham um papel importante na espermatogênese (Moro et al., 2000; Lin et al., 2001). Se situam nas regiões AZFc e AZFb respectivamente. A maioria das microdeleções observadas nos pacientes se produzem na região AZFc e se associam às patologias descritas anteriormente. Os homens com deleções maiores nas regiões AZFa y AZFb têm normalmente defeitos espermáticos como o bloqueio de células germinais (bloqueio de maturação) ou a síndrome de células de Sertoli “only” e são normalmente azoospérmicos (Vogt et al., 1996).
As mutações que impedem totalmente a função do receptor de andrógenos (AR) originam o que está definido como síndrome de feminilização testicular ou síndrome de insensibilidade androgênica completa, onde um indivíduo 46XY está completamente feminizado ao nascer (Loy et al., 2001). Além disso, determinadas mutações pontuais podem resultar em uma falha de espermatogênese (Yong et al., 2000). Os aumentos de longitude da região de repetição de trinucleotídeos (CAG), localizada no éxon 1 do gene AR, que codifica um tramo de glutaminas no domínio da ativação da transcrição do AR, se relaciona diretamente com um elevado risco de apresentar uma espermatogênese deficiente ou uma menor masculinização (Loy et al., 2001). Recentemente, em um estudo de homens com falha de espermatogênese bem documentada (Mifsud et al., 2001), se observou que homens com 26 ou mais repetições de CAG em seu AR tem 7 vezes mais possibilidades de serem azospérmicos que aqueles com menos de 26 repetições. Estes achados são compatíveis com o fenótipo da Doença de Kennedy, uma alteração neuromuscular degenerativa ligada ao cromossomo X, causada por mais de 40 repetições CAG no gene AR onde os indivíduos são inférteis devido a uma atrofia testicular e a uma oligozoospermia importante (La Spada et al., 2002). A chamada expansão CAG nesta patologia e em outras relacionadas com o mesmo fenômeno está associada com a “antecipação”, o que significa que a idade em que a doença se manifesta diminui nas sucessivas gerações. Este fenômeno pode ser explicado pela instabilidade das repetições anormalmente expandidas, o que as torna mais susceptíveis de produzir extensões ainda maiores por cada cruzamento desigual ou por adições extra de repetições em cada cadeia simples pela DNA polimerase durante a replicação meiótica do DNA nas linhas germinais masculina e feminina. Assim, o fenômeno da antecipação deve ser considerado, já que o ICSI com espermatozóides de homens com 15 a 28 repetições CAG no seu AR (que estão dentro da normalidade), as transmite de maneira estável aos seus filhos. Não obstante, para homens inférteis com maior longitude de repetições (29 a 39), o grau de instabilidade da seqüência CAG no espermatozóide não é conhecido, de maneira que se desconhece a possibilidade nestes homens com maior número de repetições, de transmitir alelos AR portadores da doença a suas filhas, que causarão a Doença de Kennedy em 50% de sua segunda geração de mulheres. Independentemente das repetições CAG, substituições de aminoácidos no domínio de união ao ligando do receptor AR que implicam os resíduos 727, 798 e 886 originam problemas na espermatogênese através de um novo mecanismo. Neste caso, a patogenicidade não se transmite por uma união deficiente do ligando e sim mediante uma interação defeituosa entre proteínas do domínio receptor com as encarregadas de mediar o processo de ativação da transcrição (Yong et al., 2000).
ASPECTOS GENÉTICOS DA INFERTILIDADE MASCULINA
O avanço do conhecimento genético diminuiu o emprego do termo “infertilidade idiopática” para classificar aqueles homens estéreis nos quais a causa de sua infertilidades é desconhecida, já que muitos fatores genéticos adquirem maior protagonismo em seu fenótipo. Além dos diretamente relacionados com a espermatogênese existem outros genes implicados na infertilidade masculina e que incluem fundamentalmente as alterações genéticas associadas a alterações na ação ou na secreção das gonadotrofinas ou no nexo genético entre a fibrose cística e a ausência bilateral congênita dos condutos deferentes (Huyn et al., 2002).
ALTERAÇÕES GENÉTICAS RELACIONADAS COM A DEFICIÊNCIA DA ATIVIDADE GONADOTRÓFICA
Estas alterações geralmente caem em duas amplas categorias: 1) aquelas relacionadas com a secreção ou com a ação do GnRH hipotalâmico, 2) Alterações primárias na secreção e na ação hipofisária de LH e FSH.
A primeira está relacionada com os sintomas comuns associados ao hipogonadismo hipogonadotrópico (HH) idiopático que incluem criptorquidia, ausência de desenvolvimento puberal, infantilismo sexual e diversos graus de hipospádias e alterações na descida dos testículos. Existe uma ampla gama de apresentações clínicas do HH dependendo do grau de ausência do GnRH, os casos mais severos manifestam-se com os sintomas anteriormente mencionados e os mais leves com desenvolvimento sexual incompleto cuja variabilidade será proporcional aos níveis de GnRH.
Geneticamente, o HH é uma alteração heterogênea em que um terço dos indivíduos possuem historia familiar positiva. Dentro deste grupo, aproximadamente 20% tem um padrão de herança ligada ao cromossomo X (Waldstreicher et al., 1996). Através do mapeamento genético, a variante ligada ao cromossomo X da síndrome de Kallman foi associada a uma mutação no gene KALIG - 1 de localização Xp22.3 (Hardelin et al., 1993). Se sugere que a proteína codificada por este gene regula a migração dos neurônios produtores de GnRH y olfativos e seu desenvolvimento.
A segunda categoria consiste em um conjunto de mutações genéticas que inativam as sub-unidades Beta do FSH e do LH e dos genes que codificam seus receptores. Os indivíduos apresentam hipogonadismo e hipoplasia das células de Leydig (Bhasin et al., 2000).
ASSOCIAÇÃO GENÉTICA ENTRE A FIBROSE CÍSTICA (FC) E A AUSÊNCIA BILATERAL CONGÊNITA DOS CONDUTOS DEFERENTES (ABCCD)
Está descrito que de 25% ou mais dos pacientes inférteis apresentam azoospermia, nos quais 1/3 é obstrutiva. Dos assim diagnosticados, 25% apresentam ABCCD (prevalência em 2% dos homens inférteis) (Patricio & Leonard, 2000).
As mutações em ambos os alelos do gene regulador da condutância transmembrana da fibrose cística (RTFC) resultam normalmente em ABCCD com uma alteração variável da função respiratória normal (Chillon et al., 1995). Para aqueles homens inférteis com ABCCD, é necessário o estudo da mulher se o casal está planejando submeter-se a um tratamento de reprodução assistida. Ainda que estejam identificados mais de 800 mutações do gene RTFC, a maioria dos laboratórios oferece estudos para as mutações mais freqüentes, incluindo a deleção de três pares de bases no éxon 10 (delta F508) e substituições únicas de bases R117H, G542X, N1303K, G551D, V520F, R553X e A455F. Se a mulher é portadora de mutação do RTFC, então a prole procedente de reprodução assistida com espermatozóides testiculares teria a mesma possibilidade de ter portadores heterozigotos e afetados depois do diagnóstico pré-natal. A opção do diagnóstico pré-implantacional (PGD) da FC para determinar o estado genético dos embriões destes casais é, hoje em dia, uma opção viável (Verlinsky et al., 1999). Mediante esta técnica, somente os portadores heterozigotos seriam transferidos ao útero materno e assim dar a oportunidades aos casais de iniciar a gestação com a segurança de que o concepto não estará afetado pela FC. O aconselhamento genético é também parte importante do processo de decisão já que diferentes combinações de mutações RTFQ produzem amplo espectro de fenótipos que variam desde severo e moderado até a ausência de sintomas.
BIOLOGIA MOLECULAR DAS CÉLULAS ESPERMÁTICAS: ANÁLISE DE DNA POR CITOMETRIA DE FLUXO
O estudo do DNA necessita para ser levado a cabo adequadamente, a utilização de moléculas fluorescentes que se unam a ele de forma eficaz e relativamente reversível (Gledhill et al., 1990). Segundo o tipo de molécula e o tipo de vínculo estabelecido com o DNA, os fluorocromos podem ser divididos em: intercalantes da dupla hélice de DNA, como o laranja de acridina (Golan et al., 1997; Samocha-Bone et al., 1998), o iodeto de propídio e a elipticina, todos eles altamente mutagênicos, dado que sua união ocorre entre os nucleotídeos. O segundo grupo é formado por moléculas que se unem externamente ao DNA, de forma que sua mutagenicidade é muito inferior, como o Hoechst 33342 (Fugger et al., 1998), a mitramicina e a cromomicina A3 (Bianchi et al., 1996). Finalmente, outras formas de união ao DNA seriam as do DAPI (diamidino-2fenilindol) (Hacker-Klom et al., 1985) e da 7-amino-actinomicina D.
EMPACOTAMENTO E INTEGRIDADE DO DNA
Alguns estudos hipotetizam que uma falha na condensação nuclear ou na resposta à descondensação induzida podem ser causas de desordens no desenvolvimento do pronúcleo masculino (Golan et al., 1997), e podem ser estudados pelo ensaio da estrutura da cromatina espermática (SCSA).
Este ensaio consiste na indução por calor ou condições ácidas in vitro e o estudo de sua reação como preditor de uma união anormal das pontes de dissulfeto da protamina e da condensação nuclear, o que poderia ser importante para diagnosticar o potencial fértil em reprodução assistida (Spanò et al., 2000). Também foram relacionadas as alterações neste aspecto com abortos de repetição (Drudy et al., 1996).
Alguns autores propuseram que estes testes deveriam estar incluídos no espermograma de rotina (Sukcharoen, 1995; Samocha-Bone et al., 1998).
A citometria de fluxo se mostra como técnica válida para medir o estado de empacotamento do DNA in vivo, assim como sua capacidade para ser descondensado artificialmente in vitro.
A base dos estudos de descondensação é a acessibilidade dos diferentes fluorocromos marcadores de DNA de aceder a este, de maneira que a acessibilidade é maior quanto mais descondensado se encontre e, como conseqüência, menor quanto mais condensado de encontre o DNA. Portanto, nos estudos realizados com citometria de fluxo, o incremento da fluorescência emitida com valor maior que o normal é sintoma de hipocondensação, enquanto que a diminuição da fluorescência é sintoma de hipercondensação. Os dois casos mostram situações possivelmente patológicas relacionadas ao empacotamento anormal da cromatina e possivelmente relacionadas à infertilidade.
Por outro lado, um DNA “saudável” deve responder adequadamente à condensação induzida, mimetizando assim a situação do espermatozóide ao entrar no oócito.
Outro aspecto a ser considerado nos estudos do DNA se refere à integridade e à presença de outras rupturas na cadeia. A forma de estudar esta situação por citometria de fluxo requer a utilização de uma técnica de biologia molecular chamada TUNEL (Gorzyca et al., 1993; Sun et al., 1997), graças a esta técnica se adicionam, por parte de uma transferase terminal específica exógena, nucleotídeos marcados com biotina em cada final 3’-OH da cadeia. Posteriormente, se unirá uma adivina covalentemente a um fluorocromo de maneira que uma elevada fluorescência refletirá o número diretamente proporcional de rupturas. Este sinal pode também ser medido por um citômetro de fluxo, sendo indetectável quando não existam rupturas de DNA, já que somente existe um final 3’-OH.
A presença de DNA descondensado em porcentagem elevada de espermatozóides pode ser especialmente relevante na injeção intracitoplasmática, já que os critérios de seleção dos espermatozóides estão baseados na morfologia e não existe forma de conhecer a situação de seu DNA in vivo, somente podendo-se trabalhar com probabilidades.
Cabe ressaltar que diversos estudos epidemiológicos foram realizados por diferentes grupos utilizando esta metodologia para examinar as influências que os tóxicos ambientais como o estireno e os pesticidas, hábitos tóxicos como o tabaco, as condições laborais e os tratamentos anti-tumorais (Spanò et al., 1998; Potts et al., 1999) têm sobre a espermatogênese.
Recentemente, realizaram-se estudos utilizando estes sistemas nos diferentes métodos de laboratório, empregados para a capacitação e congelação de sêmen. Os resultados revelaram incremento nas porcentagens de anomalias do DNA (tanto na desintegração in vitro como em sua integridade) após a criopreservação (Spanò et al., 1999; Pasteur et al., 1991) e após a centrifugação em gradientes de Percoll (Zini et al., 1999), ainda que outros autores tenham descrito melhora após a capacitação das amostras com swim-up (Spanò et al., 1999) e também com a filtração através de fibra de vidro e gradientes (Larson et al., 1999).
Finalmente, outros trabalhos tentaram estabelecer o estudo da apoptose no sêmen em bioensaio, concluindo que aproximadamente 20% dos espermatozóides ejaculados são apoptóticos e que a concentração de espermatozóides é menor naqueles que apresentam maior incidência deste fenômeno, sugerindo que a detecção da apoptose nas mostras de sêmen poderia ser método útil na determinação da qualidade seminal (Oosterhuis et al., 2000).
Atualmente existem diversos testes para medir a ruptura ou fragmentação do DNA do espermatozóide. Entre eles está o teste de dispersão da cromatina (SCD) (Halosperm ®) que se baseia na descondensação induzida do núcleo do espermatozóide.
Nosso grupo estudou a relevância deste fenômeno analisando as mesmas amostras de sêmen que se utilizaram nos tratamentos de reprodução assistida. Por um lado estudou-se a correlação entre os parâmetros seminais básicos e por outro a correlação com o sucesso dos tratamentos de IIU e FIV/ICSI.
A fragmentação do DNA se correlaciona negativamente com a mobilidade espermática, ainda que o processo de capacitação por swim-up não mostra melhora significativa na integridade do DNA.
Tanto nos tratamentos de IIU como nos de FIV não se observaram diferenças nas médias de fragmentação de DNA entre aquelas pacientes que conseguiram gravidez e as que não. Não obstante quando nos aprofundamos nos dados dos ciclos de FIV/ICSI observamos relação inversa entre a fragmentação do DNA e a taxa de fecundação, a sincronia pronuclear e a qualidade embrionária em dia 5 de desenvolvimento embrionário.
O estudo da fragmentação do DNA pode ser de especial interesse em pacientes com má qualidade embrionária em ciclos de FIV/ICSI como fator de causalidade. (Muriel et al., 2006)
ESTUDO DA ESPERMATOGÊNESE
Existem suficientes estudos utilizando a citometria de fluxo para comparar seus resultados com os obtidos por determinações histopatológicas analisando a espermatogênese, demonstrando tanto sua utilidade, como os benefícios em termos de reprodutibilidade, sensibilidade e rapidez. Os resultados mostraram boa correlação com os estudos anatomopatológicos (Mallidis & Baker., 1994; Kostakopoulos et al., 1997), demonstrando que estas técnicas podem ser empregadas como substituto da histologia clássica no estudo de rotina das biopsias testiculares.
O estudo do funcionamento da espermatogênese pode ser facilmente realizado em biópsias ou em ejaculado de homens portadores de patologias como a varicocele (Bonnanni et al., 1997), lesões graves da espinha dorsal, testículos não descendidos, obstruções anatômicas ou funcionais do trato genital (Hirsch et al., 1994), para a predição da recuperação de espermatozóides na eletroejaculação (Hellstrom et al., 1989), no impacto dos tratamentos em pacientes com câncer e na predição da espermatogênese pós-câncer (Fossa et al., 1994).
O diagnóstico por este método é tão sensível como a biópsia diagnosticada por patologista e ainda mais representativa, objetiva e rápida (em até 90 minutos), sendo especialmente importante em azoospérmicos nos quais, com a utilização de aspirados testiculares obtidos com agulha e seringa se podem detectar focos espermatogênicos nas gônadas de forma pouco invasiva (Bonnanni et al., 1997).
Estas provas podem ser realizadas utilizando tintura de DNA, que revela a ploidía do sêmen ou das células que compõem o testículo, após desagregação com métodos mecânicos ou enzimáticos (Garrido et al., 2001).
Assim se consegue obter diferentes picos ao representar a fluorescência em histograma, onde cada um deles representa células de determinadas características: sub-haplóides, haplóides condensadas, diplóides e inclusive tetraplóides, através de pequenas modificações no equipamento, diminuindo a voltagem do fotomultiplicador para “deslocar” o histograma para a esquerda e permitir alcançar sua zona (Figura 1) (Garrido et al., 2002).
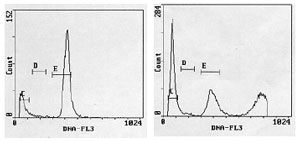
Figura 1. Histogramas de DNA de sêmen humano normal e sua correspondente distribuição de freqüências acumuladas (dados dos autores). No eixo X se representa o conteúdo relativo de DNA (dividido em 256 canais) e no eixo Y, o número de células por canal. As populações achadas foram: (C) região sub-haplóide < n (debris); (D) pico n de espermatozóides maduros haplóides ou espermátides; (E) pico 2n de espermatozóides diplóides, (F) pico >2n inclui células diplóides na fase de síntese de DNA, e tetraplóides (4n). 1) Mostra pós-capacitação, como controle haplóide 2) Controle diplóide: células sanguíneas, 3) Ejaculado com células redondas, leucócitos, determinado por tintura de Giemsa, 4) Biópsia de paciente azoospérmico, onde não foram achados espermatozóides, demonstrando-se uma existência mínima de espermátides, células diplóides e células tetraplóides.
A citometria de fluxo baseada na análise do DNA pode inclusive ser aplicada com a intenção de separar diferentes sub-populações celulares para caracterizar etapas celulares molecularmente diferentes da espermatogênese depois de realizar o “cell sorting” sendo capaz de isolar os picos anteriormente descritos de células haplóides, incluindo espermátides redondas e alargadas, diplóides e tetraplóides, considerando tanto o conteúdo do DNA como seu empacotamento (Mays- Hoopes et al., 1995, Hacker-Klom et al., 1999). É também possível comprovar a pureza das populações separadas por outros métodos (Blanchard et al., 1991).
Esta técnica poderia converter-se no tratamento de eleição em pacientes azoospérmicos secretores com deficiências extremas na espermatogênese, onde algumas áreas do testículo ainda conservam parte da espermatogênese, incluindo pacientes com “Sertoli cell only”, criptorquidias, atrofias testiculares, bloqueio de maturação espermática e síndrome de Klinefelter, permitindo-nos analisar e separar grande quantidade de células rapidamente para criar populações enriquecidas em células haplóides para serem microinjetadas, sempre que se possa utilizar marcador adequado para o DNA que não prejudique a viabilidade dos espermatozóides nem ponha em perigo a integridade da descendência.
A relação entre análise do DNA e qualidade do sêmen e infertilidade também está descrita (Hacker-Klom et al., 1999) e se concluiu que alta concentração de debris celulares, > 5% de espermatozóides diplóides ou cromatina espermática mal condensada estão relacionados com fertilidade alterada.
CONCLUSÕES
Graças às atuais técnicas diagnósticas de biologia molecular disponíveis para o estudo da espermatogênese, foram detectadas alterações neste processo que até então haviam passado despercebidas.
Estas alterações encontradas em nível molecular são a explicações do estado de infertilidade de importante número de homens.
Ainda, do conhecimento destas técnicas, se puderam desenvolver-se ou desenvolver-se-ão os tratamentos paliativos ou de reprodução assistida necessários para que os casais em que o homem esteja afetado possam ter descendência sem correr riscos.
O diagnóstico pré-implantacional é hoje em dia uma destas poderosas ferramentas. Entretanto, ainda são necessários grandes esforços para caracterizar por completo as origens das diferentes causas da infertilidade masculina para a qual, sem dúvida, as técnicas de biologia molecular terão papel decisivo.
5. Chandley A.C. Genetic contribution to male infertility. Hum Reprod., 13(Suppl 3): 76-83, 1998.
12. Golan R., Shochat L. et al
49. Verlinsky Y. & Kuliey A. Preimplantation Genetic Diagnosis. Reprod Med Rev., 7: 1-10, 1999.