JBRA Assist. Reprod 2007;11(3):9-15
ARTIGO ORIGINAL
doi: 10.5935/1518-0557.2007.11.3.02
Spontaneous and Calcium Ionophoreinduced Acrosome Reaction in Human Sperm: Roles of Capacitation Time and Conditions, Plasma Membrane Translocation of Phosphatidylserine and Protein Tyrosine Phosphorylation
Reação Acrossômica Espontânea e Induzida por Cálcio Ionóforo em Espermatozóides Humanos: Papel do Tempo e Condições de Capacitação, Translocação da Fosfatidilserina na Membrana Plasmática e Fosforilação da Tirosina
1Conceber - Centro de Medicina Reprodutiva, Curitiba, Brasil
2Reproductive Biology Unit, Department of Obstetrics and Gynecology, University of Stellenbosch, Cape Town South Africa
3To whom correspondence should be addressed at: Conceber - Centro de Medicina Reprodutiva, Av. República Argentina, 210/17° andar, 80240-210, Curitiba, Brasil - E-mail: alessandro@ clinicaconceber.com.br
4The Jones Institute for Reproductive Medicine, Department of Obstetrics and Gynecology, Eastern Virginia Medical School
ABSTRACT
The objective of these studies was to assess the relationships between capacitation time and conditions, plasma membrane translocation of phosphatidylserine (PS) and protein tyrosine phosphorylation, and the spontaneous and agonist-induced acrosome reaction of human spermatozoa. Purified populations of highly motile sperm were incubated in human tubal fluid (HTF) supplemented with varying doses of human serum albumin (HSA) and examined at 0, 3, 6, 12 and 24 hours. Acrosome reaction of live sperm was detected by fluorescein isothiocyanate-labelled Pisum Sativum Agglutinin/Hoechst; annexin V binding was used to monitor PS translocation; tyrosine phosphorylation was examined by immunoblotting. The percentage of live, acrosome reacted sperm and the percentage of live sperm depicting PS translocation increased significantly over time, both under spontaneousand calcium ionophoretreated conditions. Under spontaneous conditions there was a significant positive correlation between the percentage of live, acrosome reacted sperm and the percentage of live sperm with PS translocation. Tyrosine phosphorylation increased over time and with higher doses of HSA, and was associated with increased PS externalization. Taken together, these results suggest that the spontaneous acrosome reaction of human spermatozoa incubated under conditions that support capacitation is time dependent, is positively associated with changes in plasma membrane lipid distribution resulting in externalization of PS, and is accompanied by tyrosine phosphorylation. The responses to the agonistic effect of a calcium ionophore appear to result in different membrane dynamics.
Key words: acrosome reaction/capacitation/phosphatidylserine translocation/tyrosine phosphorylation
RESUMO
O objetivo desse estudo foi avaliar a relação entre a reação acrossômica espontânea e induzida dos espermatozóides humanos considerando o tempo e as condições, translocação da fosfatidilserina (FS) e a foforilação da tirosina. Amostras processadas de sêmen com alta motilidade foram incubadas com human tubal fluid (HTF) suplementado com variações de doses de human serum albumine (HSA) e examinas com 0, 3, 6, 12 e 24 horas. A reação acrossômica em espermatozóides vivos foi detectada por fluorescein isothiocyanate-labelled Pisum Sativum Agglutinin/Hoechst; anexina V foi usada para monitorst a translocação da FS; a fosforilação da tirosina foi examinada por immunoblotting. A porcentagem de vivos e a reação acrossômica dos espermatozóides representam o aumento da translocação da FS com maior rapidez, nas duas condições, espontânea e induzida. Em condições espontâneas houve uma positiva relação entre a porcentagem de vivos, a reação acrossômica e a porcentagem de vivos com translocação da PS. O aumento da fosforilação da tirosina com altas doses de HSA, foi associado com o aumento da liberação da FS. Associados, esses resultados sugerem que a reação acrossômica espontânea de espermatozóides humanos incubados em condições que propiciem a capacitação dependente do tempo, é ligada positivamente na distribuição de lipídios na membrana plasmática resultando na liberação da FS, acompanhado pela fosforilação da tirosina. A resposta para o efeito agonista do ionóforo de cálcio aparece nos resultados em diferentes dinâmicas da membrana.
Palavras-chaves: reação acrossômica/capacitação/translocação da fosfatidilserina/fosforilação da tirosina
INTRODUCTION
Human spermatozoa must undergo the process of capacitation in order to respond to the physiologic stimuli that trigger the acrosome reaction, a prerequisite for successful fertilization. Such agonists include follicular fluid, the steroid progesterone and the zona pellucida (Tesarik, 1985; Cross et al., 1988; Thomas and Meizel, 1989; Kopf, 1990; Blackmore et al., 1990, 5; Lee et al., 1992; Yanagimachi, 1994; Franken et al., 1996; Florman et al., 1998).
There is also a variety of pharmacological stimuli reported to trigger the acrosome reaction, either by driving extracellular calcium into the sperm cells (such as calcium ionophores) or by acting on intracellular second messengers involved in cascade reactions that couple the calcium entry with the activation of the effectors of the exocytotic response, such as pentoxifylline and others (Zaneveld et al., 1991; Cummins et al., 1991; Tesarik et al., 1993; Nassar et al., 1998).
Capacitation is a priming process that programmes the spermatozoa to arrive at the site of fertilization in a timely fashion and to rapidly undergo the acrosome reaction upon making contact with specific components of the zona pellucida (de Lamirande et al., 1997; Aitken, 1997). During capacitation, sperm undergo a variety of metabolic, functional and membrane structure modifications, eventually preparing the cell for membrane fusion and acrosomal exocytosis. In vitro studies suggest that some of these changes include alterations in sperm energy metabolism, an increase in sperm membrane calcium permeability leading to a rise in intracellular calcium levels, an efflux of cholesterol from the plasma membrane to a protein acceptor decreasing the cholesterol-phospholipid ratio, and a rise in the intracellular pH (Langlais and Roberts, 1985; Yanagimachi, 1994; Visconti et al., 1998; Cross, 1998).
Recent studies have established an association between capacitation and phosphorylation on tyrosine residues of various proteins in human spermatozoa (Aitken et al., 1995; Carrera et al., 1996; Leclerc et al., 1996; Luconi et al., 1996; Visconti and Kopf, 1998; Osheroff et al., 1999). The increase in tyrosine phosphorylation has been related to an increase in cAMP generation (Visconti et al., 1995) and also to changes in the redox status of the cells (Aitken et al., 1995, Aitken, 1997, 1).
Addition of cholesterol and an actively maintained asymmetric transmembrane phospholipid distribution characterize the membrane of mature spermatozoa during epididymal storage. In spermatozoa, as in somatic cells, the two leaflets of the plasma membrane bilayer differ in composition (Muller et al., 1994; Nolan et al., 1995; Gadella et al., 1999). The aminophospholipids phosphatidylserine (PS) and phosphatidylethanolamine are concentrated in the inner leaflet whereas the choline phospholipids sphingomyelin and phoshatidylcholine are preferentially distributed in the outer leaflet. In somatic cells, this asymmetric distribution is maintained by the action of a variety of enzymes, including flippases, floppases and scramblases (Bevers et al., 1998). An amino phospholipid translocase (also known as flippase) transfers PS from the outer to the inner lipid leaflet. Scramblases, on the other hand, transfer the various lipid species in both directions (Gadella and Harrison, 2000).
It has been hypothesized that during capacitation, an efflux of cholesterol, phospholipid movement and enhanced calcium permeability, lead to an increase of intracellular calcium and a depletion of ATP concentration. These effects, in turn, might cause a decrease in translocase activity preparing the plasma membrane for the fusion events that characterize the acrosome reaction (Nolan et al., 1995). The cholesterol-poor, lipid symmetric plasma membrane has a destabilized inner leaflet that facilitates membrane fusion upon receiving appropriate stimuli. As a result, conditions become therefore favorable for sperm to undergo the acrosomal exocytosis (Nolan and Hammerstedt, 1997).
In the present studies we aimed to further characterize the process of acrosomal exocytosis in human spermatozoa. For this purpose, we obtained purified sperm populations of high motility from fertile men (donors) and examined the spontaneous and agonist-induced acrosome reaction and their relationships with various capacitation time and conditions, plasma membrane translocation of PS and protein tyrosine phosphorylation.
MATERIAL AND METHODS
Patients inclusion criteria and study design
These studies were performed under approval of the Institutional Review Board at Eastern Virginia Medical School. Ejaculates from fertile men participating in our artificial insemination donor program were studied. Semen specimens were collected after a 2-4 day sexual abstinence period. All men were healthy, had a normal physical examination and were non-smokers. All samples had less than 1 x 106 leukocytes/mL (peroxidase staining). In order to provide sufficient number of cells for all tests, ejaculates with a concentration of motile spermatozoa = 30 x 106/mL in the original sample were included in the study.
Sperm preparation, motion analysis and morphology assessment
After liquefaction for 30 minutes at 37oC, a basic semen analysis was performed. Sperm concentration and motion parameters (i.e., the percentage progressive motility, the percentage rapid cells defined as >50 µm/s, the average path velocity or VAP, straight [VSL] and curvilinear [VCL] velocity, and linearity) were objectively evaluated using the HTM-IVOS semen analyzer (Hamilton Thorne Research, Beverly, MA, USA, version GS 771) with fixed parameter settings (Oehninger et al., 1990; Oehninger, 1995, 37). Sperm concentration and motility readings were manually monitored and corrections were made as appropriate. Sperm concentration and motility normality thresholds followed the World Health Organization criteria (WHO, 1999). Sperm morphology was examined according to strict criteria after Diff-Quik staining (Kruger et al., 1986). The semen analysis was immediately followed by separation of the motile sperm fraction by density gradient separation.The sperm fractions with high motility were isolated using discontinuous Percoll (Sigma Chemical Co, St. Louis, MO, USA) gradient separation (90% and 40% layers) and Human Tubal Fluid (HTF, Irvine Scientific, Santa Anna, CA) as diluent. Up to 2 mL of semen was carefully placed on Percoll layers, centrifuged at 380xg for 20 min and the pellet of the 90% layer was mixed with HTF and then centrifuged at 380xg for 10 min. The supernatant was discarded, the pellet was resuspended in 1 mL of capacitating medium and the sperm concentration was adjusted to 2-10 x 106/mL depending on the experiment (see below).
Assessment of acrosome reaction of live spermatozoa
The proportion of acrosome-reacted spermatozoa from initial suspensions of 2 x 106/mL motile spermatozoa was determined with the fluorescent probe fluorescein isothiocyanate-labeled Pisum Sativum Agglutinin (FITC-PSA) (Sigma) as published earlier (Cross et al., 1986; Cummins et al., 1991; Mahony et al, 1991; Tesarik et al., 1993; Oehninger et al., 1993; Nassar et al., 1998). The supravital stain Hoechst 33258 (Sigma) was used to simultaneously assess sperm viability (Cross et al., 1986). Under capacitating conditions, the percentage of spermatozoa undergoing spontaneous acrosome reaction was evaluated. To assess the agonist-induced acrosome reaction, the calcium ionophore A23187 (Sigma) was added to the sperm suspension for 60 min at a final concentration of 5 µM from a stock solution of 10 mM ionophore in DMSO.At least 200 sperm per sample were evaluated in a blind fashion at a magnification of x 1,000 using an epifluorescent microscope equipped with phase-contrast optics (Eclipse 600, Nikon, Melville, NY, USA), and using a digital camera with a high pressure mercury lamp power supply (SPOT RT, software version 3.2, Diagnostic Instruments, Augusta, GA). The following staining patterns were evaluated as live, acrosome reacted spermatozoa: (i) distinct staining in the equatorial region occurring as an equatorial bar; (ii) no staining observed over the entire sperm surface; and (iii) patchy staining on the acrosomal region (Cross et al., 1986, 9; Oehninger et al., 1993; Franken et al., 1996).
Detection of Membrane PS Translocation
We used annexin V Cy3.18 (Annexin V-Cy3; Sigma) for detection of PS externalization with simultaneous assessment of cell viability as per manufacturer’s instructions. This technique was described previously (Duru et al., 2000, 15; Schuffner et al, 2001). Briefly, in order to differentiate between live cells with and without PS translocation and necrotic cells, we used 6-carboxyfluorescein diacetate (6-CFDA) in combination with Ann V-Cy3. By microscopy, Cy3 fluoresces more brightly than the FITC conjugate. The non-fluorescent 6-CFDA enters the cell and is converted to the green fluorescent compound 6-carboxylfluorescein (6-CF). This conversion is a function of esterases present only in living cells. Thus, no fluorescence is observed in necrotic cells.Three patterns of fluorescence are typically observed: 1. Live, normal cells that stain only with 6-CF (green, annexin V-, live cells); 2. Live cells with translocation of membrane PS that stain with both 6-CF (green) and Ann V-Cy3 (red) (annexin V+, live cells); and 3.Dead cells that stain only with Ann V-Cy3 (red, necrotic).A fifty µL aliquot of sperm suspension (10 x 106/mL) was placed on a poly-L-lysine-coated slide and stained with the 6-CFDA/Ann V-Cy3 solution. After incubation in the dark for 10 min, the slide was covered with a 24x50 mm cover slip and immediately read blindly by two investigators at a magnification of 1,000x by epifluorescence microscopy as mentioned above (excitation filter: 450-490 nm, barrier filter: 515 nm). At least one hundred spermatozoa were counted per slide. The intra-observer and inter-observer variability for this technique in our laboratory are <7% and <5%, respectively (Duru et al., 2000, 15).
Electrophoresis and protein immunobloting
Sperm proteins under different incubation conditions were prepared according to Osheroff et al., (1999). Briefly, after incubation, spermatozoa from the sperm fractions with high motility were centrifugedat 20,000 g for 2 min at room temperature and then washed with phosphate-buffered saline (PBS) at room temperature. The sperm pellet was then resuspended in sample buffer (Laemmli, 1970) without mercaptoethanol and boiled for 5 min. After centrifugation at 20,000 x g for 2 min, the supernatant was removed, 2-mercaptoethanol was added to a final concentration of 5%, the sample boiled for another 5 min, and then subjected to sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE). SDS-PAGE was performed on 10% gels according to Laemmli (1970). Electrophoresis was carried out at 200 constant voltage and was continued until the tracking dye run out of the gel after loading each lane of the gel with a sperm lysate (originally containing 1 x 106 spermatozoa).After the samples were resolved by gels the sperm protein on the gel was electrically transferred to nitrocellulose membrane by using the Trans-Blot SD Semi-Dry Transfer Cell column (Bio-Rad, Mercules, CA). Membranes were blocked with blocking buffer (5% non-fat dry milk, 0.1% Tween-20 in PBS, pH 7.4) overnight at 4 °C with shaking. The primary antibody (a monoclonal antiphosphotyrosine antibody, P3300 catalog number, Sigma) was diluted in the blocking buffer and incubated with the membranes for 1 hour at room temperature. The membrane was then washed with 0.4% Tween-20 in PBS for three times, each time 15 minutes. The membranes were then incubated with a secondary antibody (anti-mouse IgG) conjugated with Horse-Radish Peroxidase (Pierce, Rockford, IL) diluted in blocking buffer (1:3,000) for 1 hour at room temperature. After three time 15 min washes with PBS/0.3% Tween-20 and three time 5 min washes with PBS/0.1% Tween-20, the blot was developed using enhanced chemiluminescence detection with ECL kit (Amersham, NJ, USA) according to the manufacturer’s instructions.
EXPERIMENTAL DESIGN
Experiment I. In the first experiment the purified fractions of spermatozoa high motility from 5 different donors were incubated under capacitating conditions in HTF supplemented with 0.3% HSA (Irvine) for up to 24 hours at 37°C under 5% CO2 in water-saturated air in order to examine the time-dependency of the spontaneous and agonist-induced acrosome reaction and their relationship with PS translocation.
At 3, 6 and 24 hours, an aliquot of sperm suspension was removed and divided into two parts: (i) the first one was kept under the same capacitating conditions for 60 min and then analyzed for CASA, spontaneous acrosome reaction and annexin V binding assay results; (ii) the other one was treated with the calcium ionophore A23187 (5 µM) for 60 min under similar capacitating conditions and then subjected to CASA, (induced-) acrosome reaction and annexin V binding assay testing.
Experiment II. In the second experiment, we examined the time dependency of protein tyrosine phosphorylation under different capacitating conditions. In experiment IIa, the purified motile fractions from 3 different donors were incubated for up to 24 hours at 37°C under 5% CO2 in water-saturated air under 2 different capacitating conditions: (i) HTF supplemented with 0.3% HSA; and (ii) HTF supplemented with 3% HSA, in order to further assess the impact of two different doses of protein supplementation on tyrosine phosphorylation. At 3, 6, 9,12 and 24 hours an aliquot of sperm suspension was removed, washed and lysed for preparation of SDS-PAGE and protein immunoblots.
In experiment IIb, we simultaneously assessed the time dependency of PS externalization and tyrosine phosphorylation using the same capacitating conditions described in experiment I. The purified motile fractions from 3 different donors were incubated in HTF supplemented with 0.3% HSA for up to 24 hours at 37°C under 5% CO2 in water-saturated air. At 3, 6, 12 and 24 hours two aliquots of sperm suspension were removed; one was used for assessment of PS externalization by annexin V binding, and the other for protein tyrosine phosphorylation by immunoblotting.
STATISTICAL ANALYSIS
The data were analyzed by using a linear mixed effects model for repeated measures. The analyses included examination of relationships between treatment groups (spontaneous conditions and after calcium ionophore), proportion of acrosome reacted live spermatozoa, proportion of sperm depicting PS translocation (annexin V positive, live spermatozoa or greenred cells, see above), and incubation time. Analysis of variance and Pearson’s correlation coefficient were used as appropriate. Data were analyzed independently by a bio-statistician (see acknowledgement). All results are presented as mean±standard deviation. P values < 0.05 were considered significant.
RESULTS
Experiment I
Table 1 shows the changes in motion parameters and viability (mean±standard deviation) throughout the 24-hour incubation period. Motility, the percentage of rapid cells, VAP, VSL, VCL and linearity declined significantly over time (p< 0.001 for all). Values for calcium ionophore-treated conditions were significantly lower than for spontaneous conditions (p< 0.002 for all motion parameters), but the rate of decline over time was statistically the same (p> 0.1 for all). Viability declined significantly over time for both spontaneous and calcium ionophore conditions (p< 0.01) as can be observed by the increase in the percentage of dead sperm assessed by Hoechst staining.
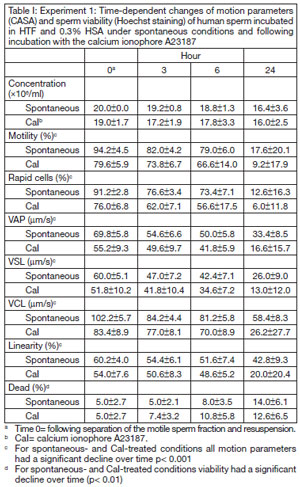
Table I. Experiment 1: Time-dependent changes of motion parameters (CASA) and sperm viability (Hoechst staining) of human sperm incubated in HTF and 0.3% HSA under spontaneous conditions and following incubation with the calcium ionophore A23187
Acrosome reaction results are shown in Figure 1. There was a time-dependent, significant increase in the percentage of live, acrosome reacted sperm throughout the 24 hour period (p< 0.002) for both spontaneous and calcium ionophore-treated conditions. Calcium ionophore responses were significantly higher that the spontaneous rates at any time point (p< 0.001).
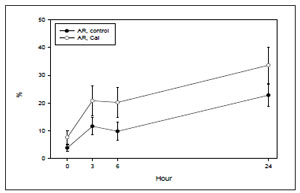
Figure 1. Experiment I: Time-dependency of the spontaneous (control) and calcium ionophore-induced (CaI) acrosome reaction of human sperm incubated in HTF and 0.3% HSA (results of 5 fertile donors).
Results of time-dependent changes of the annexin V binding assay for spontaneous (panel A) and calcium ionophore-treated conditions (panel B) are shown in Figure 2. The figure presents results of live sperm depicting PS translocation (PS translocation), live cells without PS translocation (live, normal) and dead (necrotic) cells. There was a significant increase of live spermatozoa showing PS translocation over time for both spontaneous and calcium ionophore-treated conditions (p< 0.001). However, there was no significant difference between spontaneous and calcium ionophore-treated results at any point (p> 0.1).
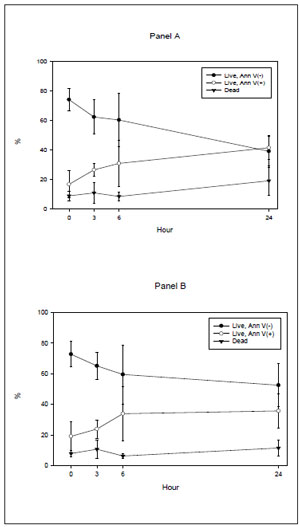
Figure 2. Experiment I: Time dependency of annexin V binding assay results of human sperm incubated in HTF and 0.3% HSA. Panel (A): spontaneous conditions; panel (B): calcium ionophore-treated conditions (Ca I) (results of same 5 fertile donors shown in figure 1).
Under spontaneous conditions, there was a significant positive correlation between the percentage of live, acrosome reacted sperm and the percentage of live sperm with PS translocation (r= 0.67, p< 0.005). However, such a correlation was not observed following calcium ionophore stimulation (r= 0.45, p= 0>0.5).
Experiment II
Experiment IIa showed that there was a timeand HSA- dependency of protein tyrosine phosphorylation profiles of highly motile spermatozoa. A representative immunoblot (panel A: 0.3% HSA; panel B: 3% HSA) is shown in figure 3. As can be observed, a subset of phosphorylated proteins of Mr (molecular mass ratio) ranging from 40,000 to 101,000 was observed at time 0 and throughout 24 hours of incubation. A time-dependent increase of phosphorylation was observed for most proteins, but particularly for the subset of proteins of Mr 95-97,000, 80-83,000 and 50-55,000. In addition, 3% HSA resulted in increased protein tyrosine phosphorylation when compared to 0.3% HSA.
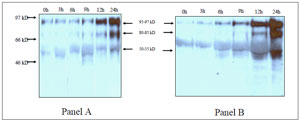
Figure 3. Experiment IIa: Time course of protein tyrosine phosphorylation of human spermatozoa incubated under capacitating conditions with two different HSA doses. Panel (A): HTF and 0.3% HSA; panel (B) HTF and 3% HSA. Numbers on the left of the figure represent markers of molecular weight (results of a representative fertile donor).
Experiment IIb showed similar temporal changes observed for PS externalization (over time increase in the percentage of live cells with PS translocation, p<0.05) that was associated with increased protein tyrosine phosphorylation. Figure 4, panel A, depicts the results of annexin V binding assay (average results of the three studied donors), whereas panel B presents a representative immunoblot from one of the donors.

Figure 4. Experiment IIb. Time course of simultaneously assessed (A) annexin V binding and (B) protein tyrosine phosphorylation by immunoblotting. Spermatozoa were incubated under capacitating conditions with HTF and 0.3% HSA (results of 3 fertile donors assessed for annexin V binding and a representative fertile donor for immunoblotting).
DISCUSSION
We hypothesized that the rate of spontaneous acrosome reaction of purified populations of highly motile spermatozoa from normal, fertile men is (i) timeand capacitation conditions-dependent, and (ii) associated with changes in membrane lipid symmetry. Our first experiment (experiment I) validated such hypothesis in that the non-stimulated acrosome reaction increased with time of incubation under capacitating conditions and acrosomal exocytosis was significantly and positively correlated with the degree of plasma membrane PS externalization. Furthermore, results of the second experiments (experiments IIa and IIb) confirmed that identical capacitation conditions resulted in a time-dependent enhancement of PS externalization and tyrosine phosphorylation of a subset of proteins, and furthermore, that such effect was HSA-dependent.
Capacitation conditions promote efflux of cholesterol from the sperm plasma membrane and destabilization of the phospholipid bilayer (Nolan and Hammerstedt, 1997; Gadella and Harrison, 2000). Here, capacitating conditions that lead to cholesterol efflux to a protein acceptor (i.e., HSA) were associated with increased annexin V binding indicating that live spermatozoa had increased membrane PS externalization. We speculate that such phenomena resulted in facilitation of membrane fusogenic effects and increased the rate of spontaneous acrosomal exocytosis.
This loss of membrane phospholipid asymmetry may be due to changes in the activities of intracellular sperm flippases and/or scramblases (Gadella and Harrison, 2000). These authors have demonstrated that bicarbonate levels, a major determinant of capacitation (Boatman and Robbins, 1991; Visconti et al., 1995) alters greatly the steady-state distribution of aminophospholipids in boar sperm as examined by labeled phospholipid probes and flow cytometric analysis (Gadella and Harrison, 2000). Such methodology obviates a possible “overestimation” of outer leaflet phospholipid distribution as a consequence of the acrosome reaction resulting in exposition of previously sequestered membranes.
We and others have also demonstrated that under other “stressinduced” conditions (i.e., cryopreservation) ram and human spermatozoa also show evidence of increased PS externalization providing indirect evidence for modification of the activity of such regulatory enzymes (Muller et al., 1999; Glander and Schaller, 1999; James et al., 1999Duru et al., 2001; Schuffner et al, 2001). The present results also extended a recent communication from our laboratory showing that prolonged incubation of human spermatozoa from fertile and infertile men at body temperature was associated with significant motility loss and membrane changes as revealed by PS translocation (Schuffner et al, 2002). Those observations as well as the results of the present studies show that although the temporal pattern of PS translocation is similar among fertile men and that there is inter-individual variability.
Our results also extended previous contributions that indicated that capacitation of human spermatozoa is associated with phosphorylation of multiple proteins at tyrosine residues (Aitken et al., 1995; Carrera et al., 1996; Leclerc et al., 1996; Emiliozzi and Fenichel, 1997; Tomes et al., 1998; Osheroff et al., 1999). Further, we have corroborated that the presence of a higher concentration of serum albumin in the culture medium results in enhanced tyrosine phosphorylation of such proteins. Osheroff et al., (1999) previously demonstrated that maximum phosphorylation of human sperm under capacitating conditions was also achieved with 3 mg/ml of bovine serum albumin and that this requirement for albumin correlated with the ability of sperm to undergo the acrosome reaction. Moreover, these authors elegantly showed that capacitation-dependent cholesterol release is associated with the activation of protein kinase A-tyrosine kinase second messenger systems resulting in protein tyrosine phosphorylation.
The ability of sperm to undergo acrosomal exocytosis in response to the agonistic effect of the calcium ionophore A23187 was also time-dependent under the capacitating conditions studied herein. However, the lack of a significant correlation with PS translocation suggests that the plasma membrane changes of the spontaneous and calcium ionophore-induced acrosome reaction may be different. These results need to be further validated by larger studies and of interest, should be compared to the effects of the physiological agonists of the acrosome reaction. Studies using either solubilized human zona pellucida and/or a biologically active recombinant human ZP3 may provide further insight into such mechanisms (Van Duin et al., 1994; Whitmarsh et al., 1996; Franken et al., 2000; Dong et al., 2001).
Acknowledgement
We acknowledge Paul Kolm, PhD, Biostatistician at Portsmouth Naval Hospital, Portsmouth, Virginia, USA, for providing independent statistical analysis.
REFERENCES
1. Aitken RJ (2000) Possible redox regulation of sperm motility activation. J Androl, 21, 491-496.
9. Cross NL (1998) Roles of cholesterol in sperm capacitation. Biol Reprod, 59, 7-11.